



ЦИКЛ ВОСПРОИЗВОДСТВА, ПОЛОВОЙ ЦИКЛ И ЕГО РЕГУЛЯЦИЯ
В. А. Середин
Воспроизводство стада – это процесс восстановления (и увеличения при расширенном воспроизводстве) поголовья стада путем размножения и выращивания более продуктивных животных, т.е. одновременно предусматривается качественное совершенствование стада, улучшение племенных и продуктивных качеств животных, направленное на решение специалистами одной из главных проблем – обеспечение населения продовольствием за счет собственных ресурсов.
В настоящее время усилия многих ученых направлены на разработку системы биотехнологии, позволяющей полнее использовать естественный репродуктивный потенциал животных. Биотехнология воспроизводства как перспективное и приоритетное направление в животноводстве развивается ускоренными темпами. Система – это совокупности элементов, находящихся между собой в определенной связи и образующих целостную функциональную структуру. Примером такой структуры является регуляция функций репродуктивной системы, от состояния управления которой зависит возможность реализации жестких параметров воспроизводства стада: обеспечение плодотворного осеменения коров в первые 1,5-2 мес. после отела и ремонтных телок в возрасте не старше 18-19 месяцев. Выдерживание таких параметров позволит фермерским молочным хозяйствам увеличить производство молока и мяса примерно на 60-70%.
Цикл воспроизводства охватывает одинаково повторяющиеся периоды определенного физиологического состояния самок. Биотехнология воспроизводства охватывает все вопросы воспроизведения животных от отела до отела, при этом важнейшим экономическим критерием, признанным во всех странах, является межотельный интервал (МОИ) или интервал между отелами (ИМО), который является интегральным показателем воспроизводительной способности коров [1].
Животные способны к воспроизведению в течение репродуктивного периода жизни, которому предшествует период полового созревания. Во время общего роста и развития организма постепенно растут и достигают зрелости половые органы, и формируется вся система ритмичного проявления половой функции. Физиологическая зрелость характеризуется завершением формирования характерного экстерьера организма и достижения им 65-70% живой массы взрослых животных данной породы и пола. Половая зрелость у телочек наступает в 5-8 – 10 мес. и зависит от породы, пола, климата, кормления, ухода и содержания, а физиологическая – в 16-18 мес., при достижении живой массы 330-350 кг. Ставится задача интенсивного выращивания телочек с обеспечением среднесуточного прироста массы тела 650-700 г, при этом повышается биосинтез стероидных гормонов и физиологическая зрелость наступает в возрасте 13-15 мес. В США применяют ранее осеменение телок в 12-месячном возрасте, при условии, что к моменту отела животное достигнет массы 450 кг. При осеменении недоразвитых самок приплод оказывается слабым и малопродуктивным (“зоотехнический брак”), общее развитие самок затормаживается.
После первой овуляции у телок половая цикличность проявляется более или менее регулярно во все сезоны года. С достижением половой зрелости у животных появляется инстинкт спаривания, а их половые железы вырабатывают способные к оплодотворению половые клетки. Плодотворное осеменение телочек должно быть обеспечено к 18-ти месячному возрасту.
В цикле воспроизводства (рис. 1) можно отметить, что проявление половой функции происходит с определенным биологическим ритмом, при этом в яичниках происходит рост и развитие одного или нескольких фолликулов, овуляция и образование желтых тел. Внешним проявлением этих изменений является течка и половая охота. Период от одной охоты или овуляции до другой называется половым циклом (ПЦ).
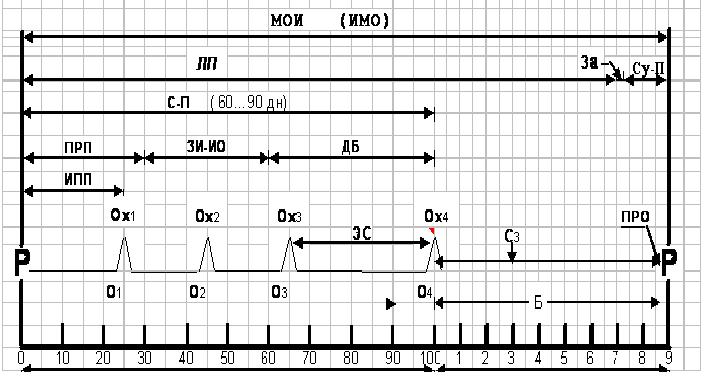
Рис.1. Цикл воспроизводства крупного рогатого скота:
МОИ - межотельный интервал (ИМО - интервал между отелами); ЛП - лактационный период; ЗА - запуск; СУ-П - сухостойный период; ПРО - постановка в родильное отделение за 10 дней до отела; С-П- сервис-период (от отела до плодотворного осеменения); ПРП - послеродовой период; ЗИ-ИО-завершение инволюции, искусственное осеменение; ДБ - дни бесплодия (С-П минус 60дн.); ИПП - индепеданс-период; Охj – охота; Оj - осеменение; Б - беременность; Сз - стельная 3 мес. (результат ректального исследования).
Оптимальный цикл воспроизводства коровы должен быть длительностью 12-13 месяцев. Экономически целесообразным является МОИ в пределах 370-380 дней. В течение жизни животного обычно наблюдается 5-7, редко 9-10 и даже 12-14 циклов воспроизводства. На протяжении цикла воспроизводства различают лактационный период (ЛП), запуск (За) и сухостойный период (Су-П) в течение последних 60 дней перед родами. В течение этого отрезка времени – от родов до родов (Р) различают сервис-период (С-П), как отрезок времени от родов до плодотворного осеменения и беременность (Б), на протяжении 9 месяцев с момента оплодотворения до очередных родов, результат ректального исследования – стельная 3 месяца (С3), при этом за 10 дней до родов, выделяется период постановки беременной самки в родильное отделение (ПРО), который определяется по появлению предвестников родов.
Продолжительность С-П наиболее целесообразна в пределах 60-90 дней. Из них 30 дней выделяется на послеродовой период (ПРП) и до 30-45 дней на завершение инволюции полового аппарата (ЗИ-ИО) и проведение осеменений животного. На графике видно, что в течение этого времени проявления половой функции наблюдается 3-4 половых цикла (ПЦ), определяемые как периоды от одной охоты и овуляции до другой (Ох1,Ох2,Ох3,Ох4), сопровождающиеся осеменением животного (О1,О2, О3,О4). Во многих странах выделяется индепеданс-период (ИПП), как период от родов до первой охоты.
Анализируя ритм половых циклов, специалист определяет нормальные интервалы ПЦ в пределах 19-21 дней и удлиненные – более 30-45 дней, связанные с ранней эмбриональной смертностью (ЭС), гибелью и рассасыванием бластоцист, эмбрионов, сопровождающихся удлинением лютеальной фазы ПЦ. Нередко удлинение интервалов между ПЦ связывают с пропуском выявления охоты у коров, особенно при неполноценных половых циклах. В.С.Шипилов (1972) считает, что 20% повторных охот связано с несвоевременным выявлением предыдущих [2].
В нормальном цикле воспроизводства, по окончании ПРП, выделяется еще 30 дней на ЗИ-ИО, поэтому дни бесплодия (ДБ) правильно должны подсчитываться путем отнятия от С-П 60 дней, что соответствует реальному состоянию воспроизводительной функции животного в каждом конкретном случае.
Репродуктивная система, обеспечивающая процесс воспроизводства, состоит из комплекса элементов (органов), функциональная активность которых определяется не только внутрисистемными сигналами, но и информацией, поступающей от других систем и из внешней среды. Вместе они составляют целостную функциональную структуру. Наибольшей надежностью обладает система представляющая собой совокупность управляемого объекта и устройства управления. Первооткрыватель кибернетики академик М.М.Завадовский, в 1924-1933гг. провел первые успешные опыты по стимуляции многоплодия у овец и коров и определил три пути скорейшего воспроизводства поголовья: 1—обеспечение многоплодия, 2 – организация уплотненных окотов или отелов, 3 – обеспечение ускоренного полового созревания. Он писал: “В силу противоречивого взаимодействия между органами развивающееся тело животного представляет собой саморегулирующуюся систему с высокой степенью устойчивости, так как регуляция, в силу связанного отношения, присуща не только организму как целому, но и каждому звену в отдельности”. М.М.Завадовский установил, что усиление функции гипофиза не только стимулирует активность половых желез (плюс-действие), но одновременно возбуждает обратную реакцию с их стороны – усиливает тормозные действия этих желез на гипофиз (минус-действие), что привело к созданию теории “плюс-минус взаимодействия” [3].
В настоящее время, в основу регулирования половой функции положен иерархический принцип: нижележащие структуры регулируются вышележащими, которые в свою очередь реагируют на изменения в нижележащих уровнях (греч. hieros–священный, arche–власть – тип отношений в сложных многоуровневых системах, характеризуемый упорядоченностью, организованностью взаимодействий между отдельными уровнями по вертикали). Иерархическая репродуктивная суперсистема характеризуется как структурой, так и функциональной дифференциацией, т.е. способностью реализации функции воспроизводства, при этом на более высоких уровнях осуществляются функции интеграции, согласования. Управление репродуктивной функцией связано с переработкой и использованием больших массивов информации, причем на нижележащих уровнях используется более детальная и конкретная информация, которая охватывает лишь отдельные аспекты функционирования системы, (например, функционирование органов-мишеней), а на более высокие уровни поступает обобщенная информация, характеризующая условия функционирования всей системы, и принимаются решения относительно системы в целом (Рис.2).
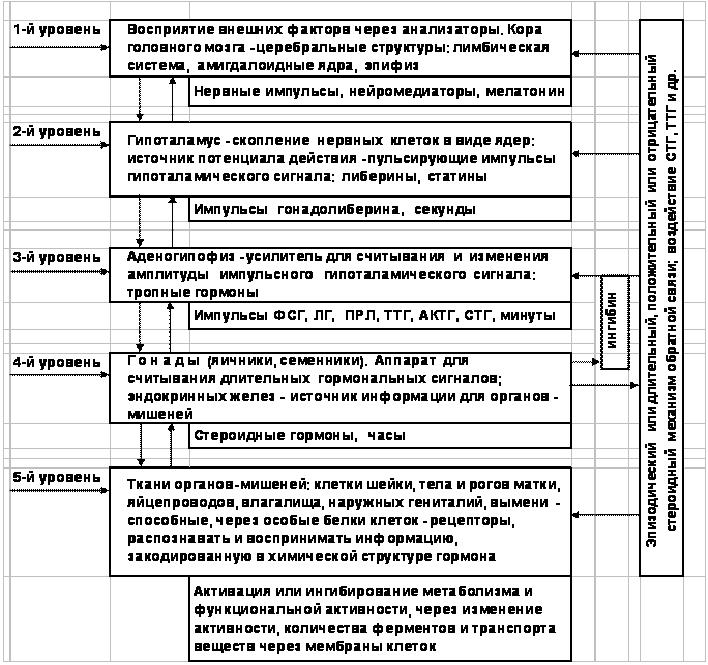
Рис.2. Уровни регуляции репродуктивной системы
Иерархическая репродуктивная система сочетается с большей или меньшей автономией нижележащих уровней по отношению к вышележащим, и в управлении используются присущие каждому уровню возможности самоорганизации.
В многоклеточном организме взаимосвязь между внешней средой, между отдельными клетками, органами и тканями, осуществляется следующими путями: нервные клетки получают сигналы, поступающие из внешней и внутренней среды, преобразуют их в форму нервного импульса и передают через синапсы, используя химические сигналы – медиаторы; эндокринная система через эндокринные железы, секретирующие в кровь гормоны, влияет на метаболизмы различных клеток-мишеней; паракринная система, когда гормоны секретируются во внеклеточное пространство и связываются с мембранными рецепторами соседних клеток (ПГФ-2а гистамин и др.); аутокринная система, когда гормон, выделенный в межклеточное пространство, связывается с мембранными рецепторами клетки, секретирующей гормон; в процессе иммунного ответа клетки синтезируют низкомолекулярные белковые соединения – цитокины, являющиеся медиаторами иммунной и воспалительной реакций, обладающие полифункциональной (плейотропной) активностью [4]. Способность продуцировать и выделять медиаторы сближает нервные клетки с эндокринными. Разница между медиаторами и гормонами заключается лишь в том, что действие первых оканчивается местом их выделения (локальность эффектов) и кратковременностью, тогда как для гормонов характерны дистантные и длительно сохраняющиеся эффекты [5]. В регуляции репродуктивной системы выделяют пять уровней.
Первый уровень регуляции репродуктивной системы
Центральная нервная система (ЦНС), состоящая из огромного количества нервных клеток (14-15 млрд. нейронов, имеющих длинные отростки – аксоны и короткие – дендриты), соединенных между собой синапсами (место контакта окончания аксона с телом или отростком другой нервной клетки). Все нейроны делятся на 3 класса: чувствительные (афферентные), которые проводят возбуждение от рецепторов в ЦНС, поэтому называются рецепторными или сенсорными; двигательные (эфферентные или эффекторные), которые передают возбуждение из ЦНС к рабочим органам–эффекторам, например, к скелетным мышцам (мотонейроны); вставочные или промежуточные – самая большая группа нейронов, осуществляющая связь между рецепторными и эффекторными нейронами.
Основными медиаторами возбуждения в ЦНС являются: ацетилхолин и нейтральные аминокислоты: глутаминовая кислота (глутамат) и аспарагиновая кислота (аспартат), которые широко распространены в ЦНС. К медиаторам, выделяющимся в тормозных синапсах, относятся кислые аминокислоты: ГАМК – гамма-аминомасляная кислота и глицин (аминоуксусная кислота). Важная роль в регулировании половой функции придается медиаторам, оказывающим как возбуждающее, так и тормозное действе на нейроны ЦНС, к ним относят катехоламины и серотонин.
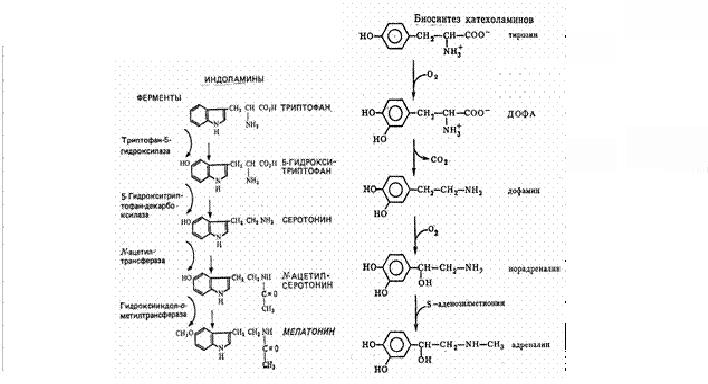
Рис 3 Биосинтез нейромедиаторов.
Катехоламины образуются из аминокислоты тирозина, а серотонин относится к индоламинам, образуется из незаменимой аминокислоты триптофана, из которой синтезируется гормон эпифиза мелатонин (Рис.3).
Выдающиеся клиницисты и физиологи конца ХIХ и начала ХХ веков С.П.Боткин, И.М.Сеченов и И.П.Павлов сформулировали положение, что животный организм связан с внешним миром постоянными связями, показали, что рефлекс – это закономерная реакция животного организма на внешний мир при посредстве нервной системы. Все акты сознательной и бессознательной жизни по способу происхождения суть рефлексы (И.М.Сеченов,1863). И.П.Павлов придавал основное значение объединяющей деятельности нервной системы и, прежде всего коры больших полушарий для всех функций организма: ”Деятельность нервной системы направляется, с одной стороны, на объединение, интеграцию работы всех частей организма, с другой – на связь организма с окружающей средой “ [6,7,8]. Наличие взаимной функциональной связи между корой головного мозга и половой системой показано классическими экспериментами И.П.Павлова, М.К.Петровой по изучению влияния кастрации на функцию высших нервных центров, а также подтверждается возможностью выработки условных рефлексов с рецепторов половых органов и изменением особенностей рефлекторных реакций в зависимости от функционального состояния организма. Н.Коренной, П.Дьяченко (1974, 1975) подробно описали клинику у животных пришедших в охоту и показали выработку условных рефлексов на окружающую обстановку и в первую очередь на оператора по искусственному осеменению [9]. В лаборатории И.П.Павлова был поставлен опыт со щенками, которых длительное время от рождения кормили только молоком. Вид и запах молока превратились в условно-рефлекторный раздражитель, всегда вызывающий слюноотделение, а другая пища – хлеб, мясо не вызывали слюноотделения, щенки не замечали предлагаемых им кусков мяса и не пытались их съесть [10].
В момент осуществления рефлекторного акта вся ЦНС представляет собой сложную мозаику, состоящую из множества возбужденных и заторможенных очагов. Более сильно возбужденный центр тормозит деятельность другого, менее возбужденного центра. Например: наложение закрутки на губу лошади вызывает сильное болевое раздражение, которое тормозит двигательные реакции животного; возбуждение центра глотания, тормозит центр вдоха и т.д.
Временное, достаточно стойкое возбуждение центра, занимающего господствующее положение в ЦНС, называют доминантой (dominare – господствую). А.А. Ухтомский (1923) приводит основные черты доминанты: повышенная возбудимость, способность к суммированию возбуждений, так как не сила возбуждения, а именно способность к длительному его увеличению под влиянием приходящих импульсов делает центр доминантным, инертным. Примером естественной гормональной доминанты является обнимательный рефлекс у самцов лягушек весной: опускание лапки в кислоту, приводит к усилению обнимательного рефлекса, и лапка не отдергивается.
Академик В.К.Милованов (1934) подчеркивает, что нужно говорить не о фолликулярной и лютеальной фазе полового цикла самки, а о смене двух доминантных состояний, определяющих специфику всей нервно-гуморальной регуляции–сексуальной (половой доминанте) и материнской (доминанта беременности). Для успешного осуществления воспроизводительных процессов половая и материнская доминанты должны своевременно сменять одна другую. В нервно-гуморальной регуляции половой функции организма ведущая и интегрирующая роль всегда остается за нервной системой[11]. На это же указывает А.П.Студенцов (1949,1950):”Половой цикл самок и половой сезон животных вообще мы расцениваем в первую очередь как реакцию организма на внешнюю среду, точнее, на условия существования, ибо сексуальные процессы у домашних животных – проявление физиологической реакции организма на действие света, пищи, тепла и на непосредственные нервные восприятия” [12]. К.М.Быков (1952) подчеркивает, что кора головного мозга, устанавливая связь всех систем организма с внешней средой, в то же время зависит от воздействия со стороны всех внутренних рецепторов организма[13]. Он провел интересные опыты по условно-рефлекторному изменению основного обмена у проводников поезда “Москва-Мурманск” при движении к Северу и обратно к Москве, как один из примеров интеграции внешнего и внутреннего, по регуляции обмена веществ, осуществляемого ЦНС.
В ЦНС имеется огромное количество коллатералей: каждый аксон дает коллатерали к целому ряду нейронов. От последних коллатерали идут к еще большему количеству нейронов и получается, что импульс пришедший в ЦНС, может иррадировать по многим направлениям ко многим центрам. В стволе головного мозга расположена ретикулярная формация, имеющая колоссальное количество связей: по ее восходящему отделу возбуждение почти диффузно распространяется к коре больших полушарий. В ЦНС афферентных путей в 4-5 раз больше чем эфферентных, поэтому к эфферентному нейрону возбуждение подходит по многим путям. Одной из особенностей прохождения возбуждения по нервным центрам является циркуляция нервных импульсов по замкнутым нейронным цепям: нервные импульсы от одного из нейронов, который пришел в возбужденное состояние, передаются как к другим нейронам, так и по коллатералям их аксонов вновь возвращаются на первый нейрон и, таким образом, возбуждение может очень длительно циркулировать в одном нервном центре до тех пор, пока не наступит утомление одного из синапсов или же активность нейронов будет блокирована торможением.
В настоящее время установлена связь лимбической системы (limbus – край, кайма–внутренняя медиальная сторона больших полушарий) с гипоталамусом и эпифизом. Шишковидная железа или эпифиз – верхний мозговой придаток расположен между буграми четверохолмия, над третьим мозговым желудочком. Соединительнотканные перегородки делят железу на дольки, внутри которых находятся пинеалоциты–крупные клетки с пузыреобразным ядром и многочисленными отростками, контактирующими с капиллярами, в них образуется мелатонин. По химической структуре он близок к серотонину, период его полужизни в крови составляет менее 10 минут. Концентрация мелатонина в организме животного определяется длительностью светового дня, т.е. это гормон связанный с фотопериодизмом. Эпифиз контролирует время наступления сезонного цикла размножения, опосредуя влияние светового режима на размножение. Удаление эпифиза превращает животное с сезонным циклом размножения в животное с постоянным размножением. Препараты: эпиталамин, синтетический мелатонин, получаемый при декофеинизации сырья, используются для регулирования сезона размножения, замедления старения репродуктивной функции. Кроме мелатонина, из эпифиза выделено много биологически активных соединений, например, аргинин – вазотоцин, активно подавляющий секрецию гонадотропных гормонов у млекопитающих.
Ведущую роль в проявлении функции размножения выполняет ЦНС, регулирующая секрецию гормонов и опосредующая их обратное действие. Внешние раздражители – триада А.П.Студенцова: корм, свет, самец – поступают через анализаторы (зрительный, обонятельный, слуховой, вкусовой, осязательный), внутренние раздражители – гормоны, сигнализируют о готовности животного к размножению [14]. Головной мозг суммирует полученные раздражения и направляет нервные импульсы гипоталамусу, где они преобразуются в гуморальные, способствующие выделению гонадотропных гормонов гипофиза. Возбужденный гипоталамус посредством ретикулярной формации индуцирует (настраивает) кору головного мозга, вызывая в ней сексуальную настроенность в поведении самки или самца, т.е. формирует половую доминанту, при этом многие функции организма мобилизуются на осуществление воспроизводства потомства [1].
Второй уровень регуляции репродуктивной системы
Гипоталамус является одним из древних подкорковых образований головного мозга и представляет собой скопление нервных клеток группами (называемых ядрами, около трех десятков), которые окружают заполненную спинномозговой жидкостью полость третьего мозгового желудочка, и проекционных путей, связывающих его с другими структурами головного и спинного мозга. В гипоталамусе имеется два типа эндокринных нейронов, которые топографически разделяются на переднюю, среднюю и заднюю группы, и синтезируют гормоны двух типов.
Крупноклеточные нейроны переднего гипоталамуса находятся в супраоптическом и паравентрикулярном ядрах, а их аксоны идут к срединному возвышению, вниз по ножке гипофиза к терминалям в задней доле гипофиза. По этим аксонам трансформируется нейросекрет, состоящий из окситоцина и вазопрессина, в заднюю долю (нейрогипофиз), где сохраняется в ней, как в резервуаре, а затем по мере надобности выделяется в кровь.
Тела мелкоклеточных нейронов образуют скопления во многих гипоталамических ядрах (преоптическом, супрахиазматическом и др.), а аксоны их обычно заканчиваются в срединном возвышении, и образовавшиеся гормоны: либерины – стимулирующие или статины – тормозящие (ингибирующие), -- через особую систему нервно-сосудистых контактов (нейрохемальные образования) попадают в кровь верхней гипофизарной артерии, прошедшую через первичное капиллярное русло срединного возвышения во вторичную капиллярную сеть передней доли гипофиза, по системе портальных сосудов.
Гипоталамические либерины (релизинг-гормоны): тиреолиберин, кортиколиберин, гонадолиберин, соматолиберин, соматостатин и др. регулируют синтез и секрецию гормонов гипофиза. Гонадолиберин (ГЛ), представляет собой цепочку из 10-аминокислот – декапептид, выделяется в пульсирующем ритме, приблизительно 1 раз в час (цирхоральный ритм секреции). Согласно современным представлениям, гипоталамический генератор пульсирующей секреции ГЛ – аркуатный осциллятор – расположен внутри гипоталамуса и представляет собой кольцевую структуру, объединяющую много сотен нейронов разных типов и функционирующую как замкнутая цепь [15]. Высота пульса ГЛ – это повышение концентрации гормона от 0-50 до 400-600 пг/мл, т.е. практически в 10 раз. Введение ГЛ в организм в пульсирующем режиме вызывало восстановление ПЦ даже у неполовозрелых животных. Структура ГЛ расшифрована и он получен синтетически. В практике стали использоваться сурфагон, гонавет, диригестран, супергестран, фертирелин и др. препараты ГЛ.
В регуляции секреции ГЛ особая роль принадлежит дофамину и серотонину, простагландинам, моноаминооксидазе (МАО), ГАМК и др. нейромедиаторам. Серотонин и гормон эпифиза мелатонин способны ингибировать выброс ГЛ и, следовательно, снижать выделение гонадотропинов (ФСГ, ЛГ) гипофизом. Дофамин – это моноамин, предшественник норадреналина, относящийся к катехоламинам, содержание которых преобладает в туберальной зоне гипоталамуса (аркуатное, вентромедиальное ядра). Норадреналин повышает продукцию ГЛ и биосинтез гонадотропинов. МАО тормозит синтез серотонина в нервных клетках и разрушает норадреналин в синапсах. Снижение уровня норадреналина приводит к снижению пульсирующего режима секреции ЛГ. ГАМК синтезируется в мозгу путем декарбоксилирования глутаминовой кислоты, в процессе обмена превращается в янтарную кислоту, которая включается в цикл Кребса, при этом высвобождается энергия, используемая в синтетических процессах мозга, в том числе и в образовании ГЛ, участвующего в регуляции половой функции [15].
Особую роль в регуляции пульсирующего ритма ГЛ играют катехолэстрогены, которые обладают сходством как со структурой эстрогена, так и со структурой катехоламина. Они замедляют метаболизм катехоламинов и, следовательно, уровень секреции ФСГ и ЛГ. Этим создается своеобразная буферная система, модулирующая функциональную активность гипоталамуса.
Третий уровень регуляции репродуктивной системы
Гипофиз, или питуитарная железа, состоит из передней доли - аденогипофиза, задней доли – нейрогипофиза и небольшой промежуточной доли, продуцирующей меланоцитстимулирующие гормоны. Это самая сложная по строению и в функциональном отношении эндокринная железа, являющаяся целой фабрикой гормонов. В аденогипофизе вырабатываются гонадотропные гормоны: ФСГ (фолликулостимулирующий – фоллитропин) и ЛГ ( лютеинизирующий – лютропин), ПРЛ – пролактин (ЛТГ – лактотропный гормон), СТГ – соматотропин (гормон роста), АКТГ – адренокортикотропный ( кортикотропин, стимулирующий надпочечник), ТТГ – тиреотропный гормон (тиреотропин, стимулирующий щитовидную железу) и др.
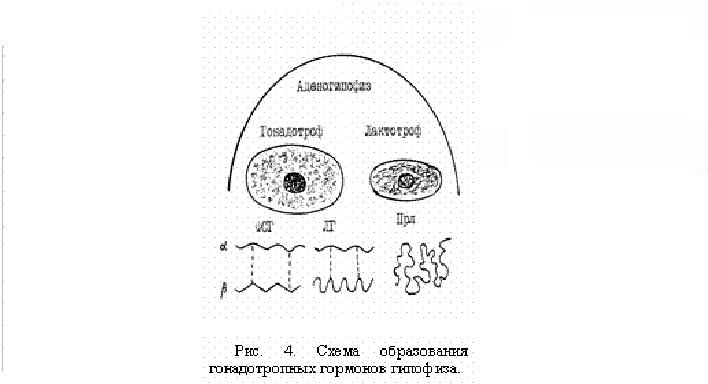
В заднюю долю гипофиза из нейронов крупноклеточных ядер переднего гипоталамуса по аксонам поступают капли нейросекрета, которые депонируются в питуицитах, а затем поступают в кровь и действуют на периферические органы. Вытяжку из задней доли гипофиза, богатую окситоцином и вазопрессином, называют питуитрином, который широко используется как окситоцический препарат, относящийся к средствам, стимулирующим мускулатуру матки. В аденогипофизе имеются клетки разных типов (Рис.4). Самые крупные – гонадотрофы (около 80% от всех клеток), в одной части этих клеток вырабатываются ФСГ и ЛГ, в другой – ТТГ, АКТГ и др. Более мелкие клетки – лактотрофы, составляют около 20% всех клеток, в них вырабатываются ПРЛ и СТГ. Молекулы ФСГ и ЛГ представляют собой гликопротеиды, имеют одинаковое строение альфа-субъединиц в молекулах из 92 аминокислотных остатков, а бета-цепи различные, в ФСГ они примерно в 1,5 раза больше альфа-цепей. Для ФСГ и ЛГ характерно наличие тонического и циклического типов секреции (Рис.5).Тоническая секреция генерируется аркуатным осциллятором гипоталамуса в цирхоральном ритме, то есть специфический сигнал в виде ГЛ выделяется приблизительно с часовым интервалом. Тоническая секреция имеется у обоих полов, поддерживает базальные уровни ФСГ и ЛГт, которые обеспечивают как развитие половых клеток (гамет), так и эндокринных элементов половых желез (гонад). Эстрогены и прогестерон по механизму обратной связи влияют на секрецию ФСГ и ЛГ как на гипоталамическом, так и на гипофизарном уровне. Клетками гранулезы фолликулов, клетками Сертоли извитых канальцев семенников вырабатывается белок ингибин, который тормозит гипофизарную секрецию ФСГ. Т1/2 ФСГ составляет примерно 150 мин, а Т1/2 ЛГ – 30 мин. Циклическая секреция существует только у самок, функционирует кратковременно – на протяжении 12-24 часов в течение каждого ПЦ, когда содержание ЛГц в крови увеличивается иногда в несколько сотен раз. Главной функцией ЛГц является индукция овуляции у самки. Центры циклической секреции располагаются в преоптической области переднего гипоталамуса. Органами – мишенями для ФСГ и ЛГ являются гонады. В яичнике самок ФСГ стимулирует рост фолликулов, находящихся в стадии антрум (полостных), на первичные фолликулы он не действует. ФСГ вызывает пролиферацию клеток гранулезы и стимулирует образование рецепторов ЛГ на их поверхности. У большинства видов выброс ЛГц не зависит от полового акта, это животные со спонтанной овуляцией, и только у кролика, кошки, хорька и верблюда циклическая секреция ЛГ индуцируется половым актом, т.е. они относятся к животным с индуцированной или рефлекторной овуляцией. М.И.Прокофьев (1979) выявил прямую зависимость между оплодотворяемостью коров и уровнем ЛГц в их крови в предовуляторный период [16].
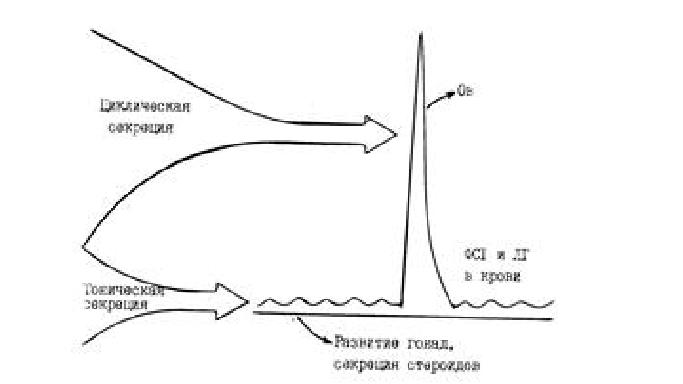
Рис 5. Схема двух различных видов секреции ФСГ и ЛГ
Гормон лактотрофов ПРЛ усиливает гормональную функцию ЖТ и активность прогестерона, влияет на метаболические процессы, оказывает трофическое воздействие на гонады и органы- мишени, стимулирует рост протоков вымени и оказывает синергичное прогестерону действие на дольчато - альвеолярную систему вымени, вместе с кортикостероидами поддерживает секрецию молока в развитом вымени. ПРЛ близок по химическому строению к СТГ, представляет собой полипептидную цепочку из 197-199 аминокислотных остатков. В отличие от ФСГ и ЛГ он не подвержен действию закона обратной связи и регулируется по короткой обратной связи гипоталамус – гипофиз методом ауторегуляции системой гипоталамического дофамина. Синтез и секрецию ПРЛ стимулируют тиреолиберин, серотонин, окситоцин, ацетилхолин, тормозит дофамин. Период полураспада ПРЛ составляет 15-20 минут.
ПРЛ остро реагирует на все виды стресса, характеризуется суточным ритмом секреции (сон – бодрствование). Ночной пик ПРЛ достигает 50% от дневной концентрации. При беременности концентрация ПРЛ возрастает в 20-40 раз (с 5-10 до 200 нг/мл). При гиперпролактинемии тормозится стероидогенез в яичниках и развитие фолликулов, например, при подсосном методе выращивания телят возникает лактационная анафродизия, при этом тормозится не только выработка ГЛ, но и тормозится гонадотропная функция гипофиза [1].
Четвертый уровень регуляции репродуктивной системы
Гонады самок – яичники (Рис.6). Это половые железы с двойной функцией: репродуктивной – в них происходит фолликулогенез и эндокринной – образуется большое количество гонадальных гормонов, главные из которых стероидные гормоны 17-бета-эстрадиол (Е2) и прогестерон (П). Менее активными являются эстрон (Е1) и эстриол (Е3). Т1/2 прогестерона в крови составляет 5 мин, эстрогенов – 0,5-1,5 часа.
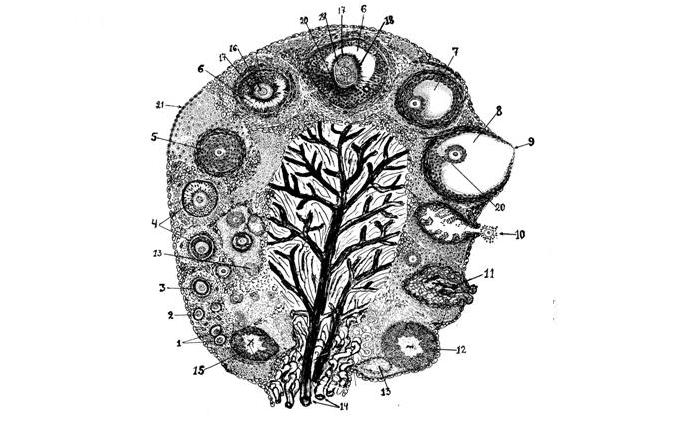
Рис. 6. Схема фоликулогенеза , овуляции и образования желтых тел в яичнике коровы: 1 – ооциты в корковом слое яичника; 2 – примордиальный фолликул; 3 – первичный фолликул; 4 – образование двухслойного фолликула; 5 – многослойный фолликул и образование теки; 6 – вторичный фолликул в стадии антрум –- образование полости с фолликулярной жидкостью;
7 –- третичный или графов фолликул; 8 – предовуляторный или доминантный фолликул перед овуляцией; 9 - стигма; 10 – овуляция – выход яйцеклетки через разорвавшуюся стенку яичника, вместе с фолликулярными клетками и жидкостью фолликула; 11 – образование геморрагического желтого тела в полости бывшего фолликула;12 – полностью сформировавшееся желтое тело; 13 – атретический фолликул; 14 – кровеносные сосуды и нервы; 15 – регрессирующее желтое тело (обратное развитие); 16 – ядро яйцевой клетки; 17 – прозрачная оболочка (зона пеллюцида); 18 – лучистая корона из фолликулярных клеток (корона радиата); 19 –- желток яйцевой клетки, равномерно распределенный в цитоплазме; 20 – яйценосный бугорок; 21 – целомический эпителий, покрывающий яичник.
У половозрелой телочки в обоих яичниках около 75-100 тысяч яйцеклеток. В процессе жизни большая часть фолликулов дегенерирует, подвергается атрезии (обратное развитие без овуляции), при этом яйцевая клетка гибнет и к 9-ти летнему возрасту коровы, их остается около 2,5 тысяч. Целостность покровного эпителия и белочной оболочки периодически нарушается во время овуляции, поэтому ткань яичника непрерывно находится в состоянии повреждения, восстановления и регенерации. Значительная часть яичника образована временными структурами, такими, как фолликулы и желтые тела (ЖТ), поэтому относительные размеры яичника постоянно меняются в течение ПЦ и цикла воспроизводства.
Зрелый фолликул (третичный или доминантный) имеет полость до 2 см в диаметре, заполненную фолликулярной жидкостью. Слой гранулезных клеток, прилегающий к базальной мембране, называется зернистым слоем. Гранулезные клетки – фолликулярные эпителиоциты, прилегающие к прозрачной оболочке ооцита, содержат микротрубочки и называются лучистой короной (Corona radiata), а группа клеток, на которой находится ооцит называется яйценосным бугорком (Рис. 7). В соединительнотканную капсулу фолликула врастают многочисленные кровеносные капилляры и она дифференцируется на два слоя: внутренний (theca interna), состоящий из текальных эндокриноцитов, прилегающих к базальной мембране фолликула, и наружный (theca externa), образованный гладкими миоцитами и волокнистой соединительной тканью.
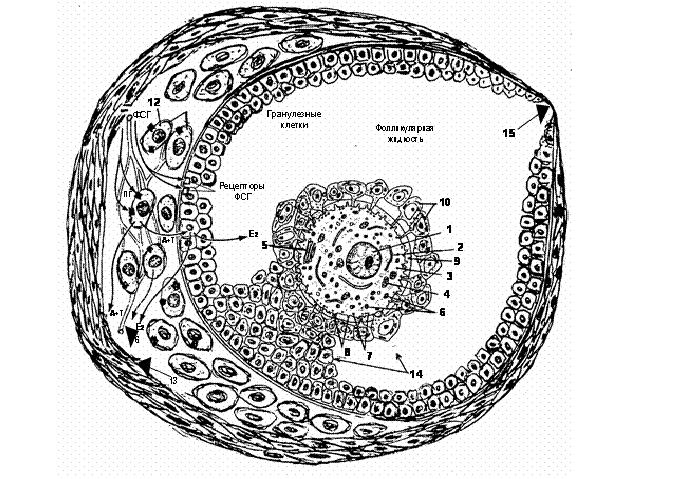
Рис. 7. Схема фолликула и действия гонадотропных гормонов гипофиза на фолликул и синтез эстрогенов: 1 – ядро яйцеклетки (ооцита); 2 – гранулярная эндоплазматическая сеть; 3 – цитоплазма яйцеклетки; 4 – митохондрия; 5 – комплекс Гольджи; 6 – желточные гранулы; 7 – кортикальные гранулы; 8 – микроворсинки; 9 – прозрачная (зона пеллюцида) оболочка яйцеклетки; 10 – фолликулярные эпителиоциты – гранулезные клетки, окружающие яйцевую клетку, образуют лучистую корону, содержат микротрубочки; 11 – базальная мембрана и прилегающие к ней гранулезные клетки, называемые зернистым слоем-гранулезой; 12 – внутренний сосудистый слой фолликула, состоит из клеток стромы – текальные эндокриноциты и богатой сети капилляров, называется тека интерна; 13 – наружный слой фолликула из клеток стромы и фиброзной соединительной ткани, называемый тека экстерна; 14 – яйценосный бугорок с яйцеклеткой; 15-стигма.
ЛГ взаимодействует с рецепторами на текальных эндокриноцитах (theca interna) и стимулирует в них выработку андрогенов – андростендиона и тестостерона (А+Т) и небольших количеств Е2. Под влиянием ФСГ в клетках гранулезы (зернистый слой) активируется система ферментов, осуществляющих ароматизацию андрогенов (А+Т) и превращение их в эстрогены, главным образом, в Е2. Концентрация 17-бета-экстрадиола в полости развивающегося фолликула в 1000 раз превышает его концентрацию в кровотоке, что влияет на созревание ооцита, дифференцировку гранулезных клеток и развитие рецепторов гонадотропных гормонов. В ответ на выделение большого количества Е2, по принципу обратной связи, центры, расположенные в преоптической области гипоталамуса, выделяют ГЛ, обеспечивающий выброс овуляторного пика ЛГц (Рис.5). Генетически ооцит блокируется на стадии диплотены мейоза на многие годы, до предовуляторного выброса ЛГц и только спустя 24 ч, после выброса ЛГц в доминантном фолликуле возобновляется мейоз, и в стадии метафазы II происходит овуляция.
ФСГ стимулирует выработку гранулезными клетками протеолитического фермента – активатора плазминогена, что разрушает основное вещество и отделяет ооцит от яйценосного бугорка, при этом рвутся плазматические мостики между ооцитом и клетками яйценосного бугорка. ЛГ стимулирует выработку PGF-2альфа и PGЕ2 в фолликулярной жидкости преовуляторного фолликула, возможно стимулирует высвобождение коллагеназ, подготавливающих фолликул к овуляции. Хотя объем фолликулярной жидкости существенно не увеличивается, внутрифолликулярное давление перед разрывом не возрастает. Установлено, что простагландины группы Е и F оказывают сильное стимулирующее влияние на миометрий, а гладкие миоциты содержат сократительные белки – актин и миозин, что возможно способствует выделению ооцита, через разрыв в области стигмы. В момент овуляции вся масса гранулезных клеток яйценосного бугорка вместе с ооцитом медленно выделяется из фолликула вместе с довольно вязкой фолликулярной жидкостью. В.Прохорова (1999) приводит 10 снимков процесса овуляции, зарегистрированного ультраэхографией, который происходит у кобыл в среднем за 42 секунды (от 5до 90 секунд) [17].
После разрыва фолликула строма яичника сокращается, стенка фолликула спадается, становится складчатой (фестончатой). Кровь, вытекающая из мелких сосудов вместе с остатками гранулезных клеток, образует сгусток (геморрагическое ЖТ) и в него врастают сосуды, направляясь вдоль тяжей клеток. Клетки ЖТ – лютеоциты происходят из двух видов клеток: из клеток внутреннего слоя теки мелкие лютеоциты (диаметром 10-20 мкм) продуцируют эстрогены и крупные (диаметром больше 25 мкм), которые происходят из гранулезных клеток и продуцируют прогестерон (П). Увеличение массы ЖТ после овуляции связано в основном с увеличением размеров лютеоцитов (гипертрофия), а не с увеличением их числа (гиперплазия). Нормальное ЖТ образуется лишь в том случае, если в фолликуле имеется достаточное число гранулезных клеток с большим числом рецепторов для ЛГ. После преовуляторного выброса ЛГц митотическая активность гранулезных клеток прекращается, поэтому увеличение массы ЖТ после овуляции связано с нарастанием размеров отдельных клеток [18].
Образование и поддержание ЖТ зависит от секреции ЛГ и ПРЛ передней долей гипофиза. Основным секреторным продуктом ЖТ является прогестерон–гормон сохраняющий беременность. Кроме него в ЖТ синтезируется два полипептидных гормона: релаксин и окситоцин, а также простагландины и простациклин. П вызывает переход слизистой оболочки матки из фазы пролиферации (вызываемой эстрогенами) в фазу секреции. Он преобразует эндометрий в прегравидное состояние – происходит усиленное разрастание маточных желез и их секреция, увеличивается содержание гликогена, ферментов, принимающих участие в образовании эмбриотрофа, необходимого для питания развивающейся бластоцисты и обеспечения ее имплантации к слизистой оболочке матки.
Пятый уровень регуляции репродуктивной функции
Ткани органов-мишеней, являются конечной точкой действия половых гормонов, вырабатываемых яичниками. В организме нет такой функции, которая не была бы подвержена контролирующему действию стероидных гормонов, поэтому они являются главными гормонами, поддерживающими постоянство внутренней среды (гомеостаз). Ткани репродуктивных органов являются специфическими органами-мишенями для половых гормонов.
Начальный этап в действии гормона на клетку-мишень – взаимодействие гормона с рецепторами клетки, имеющим центр связывания с гормоном. Для стероидных гормонов характерен внутриклеточный (цитозольный) механизм действия, так как они относятся к липофильным веществам (растворимым в жирах), поэтому способны проникать через липидный слой плазматической мембраны внутрь как клеток-мишеней, так и клеток, не являющихся объектом их биологического действия. В клетках-мишенях молекула стероидного гормона задерживается надолго – на десятки минут и даже часов, так как в цитоплазме клеток-мишеней есть рецепторы стероидов, которые связывают молекулы гормона (Рис.8).
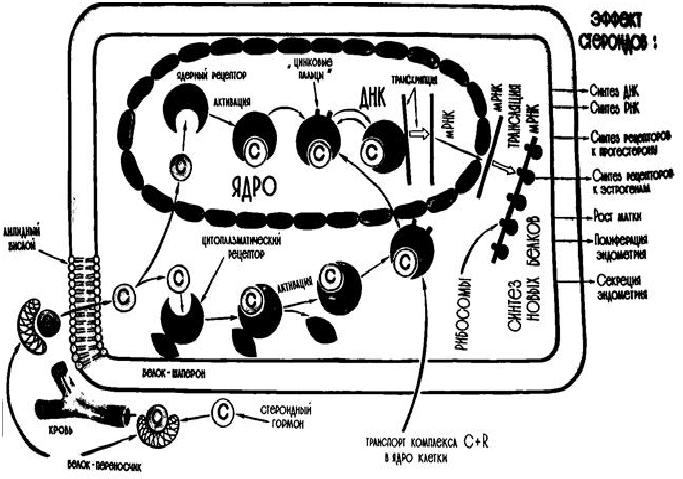
Рис.8. Механизм действия эстрогенов и других стероидов
Рецепторы стероидных гормонов – это гидрофильные олигомерные белки, которые, как правило, состоят из трех доменов. На С-концевом участке полипептидной цепи рецептора находится домен узнавания и связывания гормона. Центральная часть рецептора включает домен связывания ДНК. На N-концевом участке полипептидной цепи располагается домен, называемый вариабельной областью рецептора, отвечающий за связывание с другими белками, вместе с которыми участвует в регуляции транскрипции [4]. Концентрация рецепторов в тканях-мишенях составляет около 10-10 – 10-9 М, а количество – 5-40 тысяч молекул на клетку. Для проявления эффекта стероидного гормона необходимо, чтобы гормон-рецепторный комплекс, образовавшийся в цитоплазме, подвергся активации и проник в ядро клетки, где он регулирует транскрипцию структурных генов. Проникнув в ядро, гормон-рецепторный комплекс связывается как с ДНК, так и с белками хроматина. Индукция или репрессия синтеза белка происходит лишь спустя 1-2 часа после проникновения в клетку стероидного гормона.
Предполагается, что эстрогены индуцируют синтез свыше 50 различных белков, участвующих в проявлении физиологических эффектов эстрогенов. Они стимулируют развитие тканей, участвующих в размножении, определяют развитие вторичных половых признаков, в лютеиновой фазе ПЦ, вместе с прогестинами эпителий матки переводят из фазы пролиферации в фазу секреции эмбриотрофа, подготавливают эндометрий к имплантации бластоцисты. Совместно с PGF-2альфа увеличивают чувствительность миометрия к действию окситоцина во время родов, оказывают анаболическое действие на кости и хрящи, поддерживают нормальную структуру кожи и кровеносных сосудов. Стероидные гормоны не только регулируют функцию размножения и развития, дифференцировку органов и тканей, но и контролируют все виды обмена веществ. В настоящее время установлены закономерности половой дифференцировки мозга: во всех областях мозга имеются нейроны (около 1% всех нейронов), связывающие половые стероиды, причем распределение нейронов, в которых связывается тестостерон, очень сходно с распределением нейронов, связывающих эстрадиол, но не абсолютно идентично. В аденогипофизе 60-80% клеток поглощают Е2, в то же время в гипоталамусе только 50% нейронов могут поглощать Е2, но в 2-3 раза менее интенсивно, чем в гипофизе. Деятельность репродуктивной системы направлена на воспроизводство, т.е. на существование видов животных, при этом на всех уровнях регуляция осуществляется нервной, эндокринной, паракринной и аутокринной системами, обеспечивающих единство организма и внешней среды. В соответствии с теорией “плюс-минус взаимодействия” М.М.Завадовского (1929-1933 гг.) можно выделить следующие петли обратной связи (Рис.9):
- длинную петлю обратной связи, между гонадальными гормонами яичника и ядрами гипоталамуса; между гормонами яичника и гипофизом;
- короткую петлю между аденогипофизом и гипоталамусом; между эпифизом и нейроцитами гипоталамуса; между гипоталамусом и корой головного мозга;
- ультракороткую петлю между ядрами нейроцитов гипоталамуса.
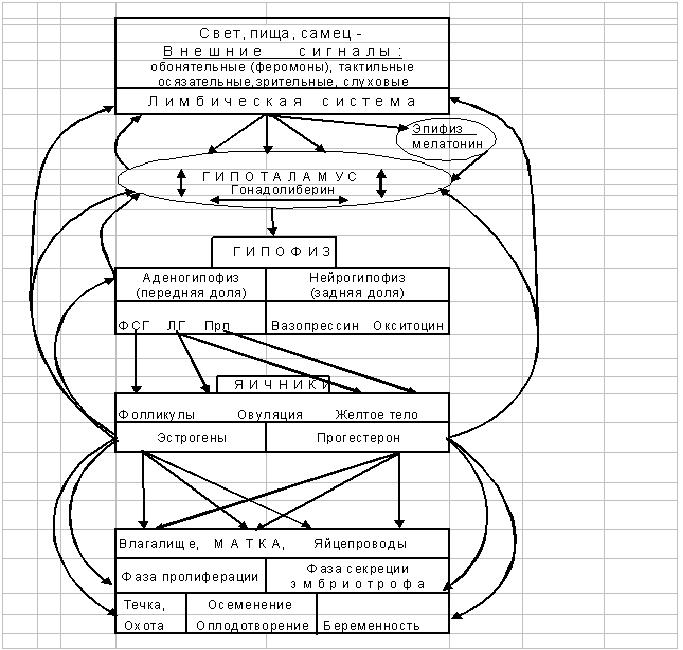
Рис. 9. Петли прямой и обратной связи в регулировании репродуктивной функции
Концепция цикла воспроизводства, а также петли прямой и обратной связи в регулировании иерархической репродуктивной системы, осуществляющиеся между различными ее уровнями, должны использоваться в практике интенсификации воспроизводительной функции животных. К настоящему времени ветеринарная наука и практика располагают достаточно эффективной и апробированной системой мероприятий, позволяющих реализовать в производственных условиях высокие потенциальные возможности репродуктивных свойств крупного рогатого скота.
С наступлением половой зрелости у телок и окончанием послеродового периода у коров возобновляются половые циклы-ПЦ. На графике цикла воспроизводства их четыре, и затем наступает беременность, и вновь повторение циклов воспроизводства до старости коровы (климактерический период), хотя практически корова не доживает до этого возраста. За всю жизнь коровы наблюдается 4-8 циклов воспроизводства и редко до 10-12, при этом каждый раз количество ПЦ может быть различным - от 1-2 до 3-5 и больше в зависимости от оплодотворяемости, имеющей полифакторную этиологию, связанную с многими паратипическими факторами, квалифицированным контролем за состоянием репродуктивной функции и безупречным физиолого-клиническим статусом стада [1]. Не случайно основоположник метода искусственного осеменения И.И.Иванов писал, что “процент оплодотворяемости прямо пропорционален квалификации и честности работника” [19].
По определению Р.В.Шорта ПЦ является лишь следствием стерильности и тем способом, с помощью которого природа гарантирует наступление очередной беременности [20]. Таким образом, биологическое значение изменений, происходящих на протяжении ПЦ, состоит в осуществлении репродуктивной функции: созревании яйцеклетки, ее оплодотворении и развитии беременности. Ритмический характер активности репродуктивной системы определяется яичником (“игра оркестра, в котором роль дирижера выполняют гонады”)[15]. Двухфазный характер связан с ростом и созреванием фолликулов, овуляцией и развитием желтого тела-ЖТ в яичнике. Длительность ПЦ в днях зависит от времени, которое необходимо для созревания и овуляции доминантного фолликула (фолликулярная фаза) и протекания стадий формирования, расцвета и регрессии желтого тела (лютеальная фаза), разделенных между собой процессом овуляции, т.е. разрывом доминантного фолликула и выходом из него зрелой яйцеклетки с окружающими ее фолликулярными клетками и жидкостью (Рис.10).
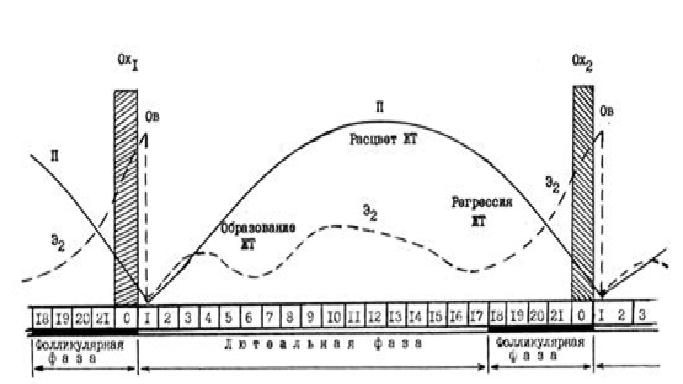
Рис.10. Длительность полового цикла в днях: зависит от времени, которое необходимо для созревания и овуляции доминантного фолликула, протекания стадий формирования, расцвета и регрессии ЖТ
Половые гормоны-сексагены (Sex-пол) выделяются в кровь не только половыми железами, но и плацентой, надпочечниками. Все сексагены образуются из холестерола (рис.11)
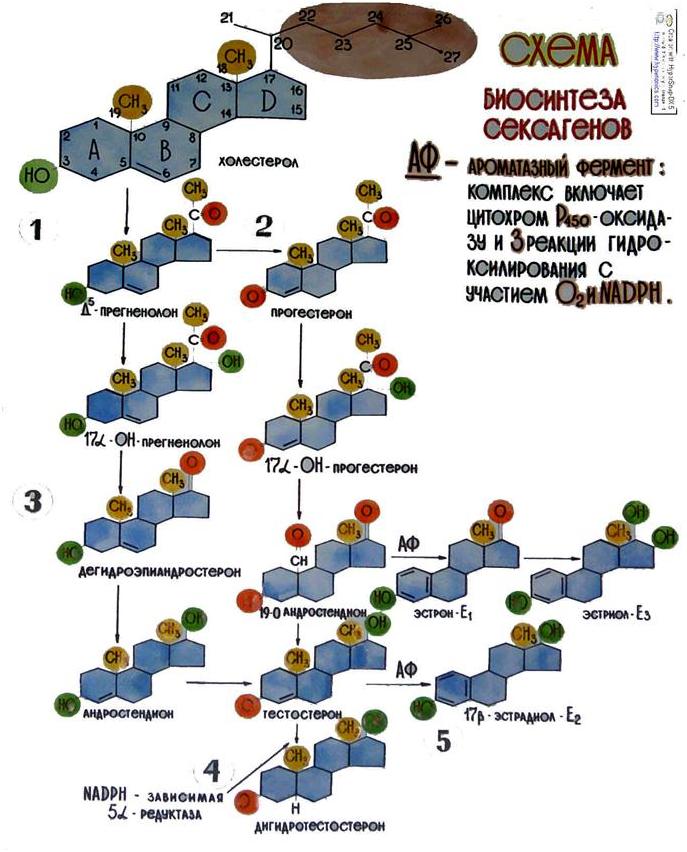
Рис.11. Биосинтез сексагенов
Сексагены являются производными циклопентанофенантренового ряда, состоят из четырех колец А, В, С и D, которые составляют скелет стероидной молекулы. Они делятся на четыре основные группы: С18- стероиды-эстрогены(от лат.oestrus- течка, греч.oistros-страстное влечение) с основными представителями - 17- бета- эстрадиол- Е2, эстрон-Е1 (фолликулин) и эстриол-Е3; С19 – стероиды-андрогены(andros, aner-мужчина), важнейшими представителями которых является дегидроандростерон, (Д А), андростендион (А) и тестостерон (Т); С21 – стероиды- гестагены, прогестины (hestatio-беременность) с основным представителем прогестероном (П) и С21 - стероиды- кортикостероиды, продуцирующиеся корой надпочечников. Сексагены являются главными гормонами, поддерживающими постоянство внутренней среды, регулируют функцию размножения и развития, дифференцировку органов и тканей, контролируют все виды обмена веществ, влияют на генетическую активность, действуют на мембранные структуры клеток, имеют многочисленные эффекты во всех органах и тканях.
В настоящее время подтверждается гипотеза о наличии двух тканевых структур, синтезирующих Е2 в развивающемся фолликуле: клетки теки интерна взаимодействуют с рецепторами ЛГ и способны как продуцировать андрогены (андростендион и тестостерон), так и ароматизировать их до Е2, при этом образующийся Е2 поступает непосредственно в кровяное русло; клетки гранулезы, взаимодействуют с рецепторами ФСГ, продуцируют прогестерон и ароматизируют поступающие к ним от теки андрогены, превращая их в Е2, который накапливается в фолликулярной жидкости и не поступает в периферический кровоток [15].
В фолликулиновой (пролиферативной) фазе ПЦ установлена положительная коррекция между уровнем Е2 в плазме крови и концентрацией рецепторов эстрогенов в эндометрии. Повышение уровня рецепторов Е2 в предовуляторном периоде сопровождается преобладанием цитозольной фракции над ядерной, а в лютеиновую – наоборот. Суммарное количество рецепторов прогестерона больше, чем эстрадиола, на протяжении всего ПЦ, за исключением начала фолликулярной фазы. Более половины рецепторов внеядерной локализации ассоциированы с цитоплазматическими структурами, в основном с шероховатым эндоплазматическим ретикулумом, который считается местом синтеза рецепторов Е2 и П. В лютеальной (секреторной) фазе количество Е2 в эндометрии снижается в два раза, а прогестерона, наоборот, возрастает в 3-4 раза [15].
Продолжительность ПЦ у коров составляет 19-21 день с отклонениями от 17 до 24 дней. Большинство исследователей за норму принимают ПЦ длительностью 18-25 дней у коров и 17-24 дней у телок, при этом доля ПЦ с нормальной продолжительностью повышается с 36,1 до 82,5% с увеличением порядкового номера цикла, т.е. с отдалением срока овуляции от времени отела и восстановления маточных структур [21,22]. ЭС колеблется в пределах 20-30% от оплодотворившихся коров и зависит от экзо- и эндогенных факторов [1].
В учении о половом цикле возникло много различных суждений, отражающих различные школы ученых, хотя не они являются определяющими, если обратить внимание на цикл воспроизводства и МОИ, которые являются интегральными экономическими показателями в скотоводстве [1,11,23,24].
Приводим описание определений ПЦ данных разными авторами.
Русский профессор Ф.Унтербергер (1802-1862) обратил внимание на цикличность половых процессов. Он писал: “У самок процесс периодического возникновения охоты зависит от внешних условий: времени года, кормления, ухода, помещения”. [23, 24].
Русский исследователь Буш предложил различать в период течки три фазы – начальную, среднюю и окончательную,-на основании клинических изменений полового аппарата и проявлений полового возбуждения. В каждой фазе описывались изменения слизистой оболочки влагалища и характера вагинальной слизи. Данные Буша легли в основу книги Крейца (1867) “Опыт полной практической инструкции для коннозаводства”. Описанный Ф.Унтербергером способ случки кобыл, применявшийся на заводе Белякова (1838), был принят под названием “русского метода” и применялся на Вюртемберском конном заводе и на немецких королевских заводах [23].
Английский зоолог из Кембриджа Уолтер Хип (Heap, 1900) ввел понятие следующих стадий ПЦ: проэструс – предтечка, эструс – течка, метэструс – стадия, после течки, когда признаки полового возбуждения сглаживаются, диэструс – стадия активности желтого тела, период относительного полового покоя, анэструс – отсутствие течки, период полового покоя. Эта терминология применяется у лабораторных грызунов, а для более крупных домашних животных и приматов она не подходит [20].
Впервые в литературу термин “ПЦ” ввел Маршал: “Под ПЦ следует понимать все многообразие структурных и функциональных изменений, происходящих в половом аппарате, эндокринной и других системах организма самки от одной половой охоты до другой (или от одной овуляции до другой). Период времени между двумя половыми охотами (или овуляциями) составляет продолжительность ПЦ” [25].
Профессор Казанского ветеринарного института имени Н.Э.Баумана, Лауреат Сталинской премии А.П.Студенцов (1949,1953), исходя из нервных процессов, возникающих в организме самок на протяжении ПЦ, предложил различать в ПЦ три стадии: возбуждения, торможения и уравновешивания. Он писал: “Под ПЦ понимается комплекс физиологических процессов в половом аппарате и во всем организме самки, протекающий от одной стадии возбуждения до другой. Ритм ПЦ, последовательность и взаимосвязь сексуальных явлений (овуляции, течки, охоты и полового возбуждения) могут быть объяснены взаимодействием нервной и гуморальной систем организма… Первичные импульсы эта гуморальная система получает от коры головного мозга”. ”ПЦ – нейрогуморальная реакция самки на факторы внешней среды, с которой организм животного составляет единство. Из внешних факторов, обуславливающих ПЦ, следует отметить три главные: 1) корм, 2) свет и 3)самец, как стимулятор сексуальной системы.”[12,14].
Академик В.К.Милованов (1934-1962) подчеркивает, что возникновение ритмической смены половых и материнских рефлексов нельзя рассматривать как смену только двух противоположных состояний самки: фолликулярной и лютеальной фаз. Такое понимание несколько ограничено, узко, так как связывает изменения в организме самки только со сменой фолликулов и желтых тел в яичнике, в то время как в нейрогуморальной регуляции половой функции организма ведущая и интегрирующая роль всегда остается за нервной системой. Поэтому нужно говорить о смене в организме самки двух доминантных состояний – сексуальной и материнской… Для успешного осуществления воспроизводительных процессов половая и материнская доминанты должны своевременно сменять одна другую [11].
В США Г.У.Солсбери и Н.Л.Ван-Демарк (1966): “Воспроизводительная система самок большинства млекопитающих характеризуется ритмическими изменениями, называемыми течковым циклом. Центральным моментом ПЦ является период течки, в течение которого самка подпускает к себе самца, а вскоре после течки из фолликула выделяется яйцо… В настоящее время часто употребляют термин фолликулярная, или эстрогенная, фаза, что бы обозначить периоды течкового цикла, в которых преобладает влияние эстрогенных гормонов. Ту часть цикла, когда развивается желтое тело и прогестерон становится господствующим гормоном, называют прогестационной или лютеальной фазой [26].
Академик В.С.Шипилов, определяет ПЦ по А.П.Студенцову и полемизируя с В.К.Миловановым (1949) за введение понятия половая доминанта заключает, что поскольку ПЦ при отсутствии беременности возникают, проявляются и угасают через определенные промежутки времени (в среднем через три недели), то мы должны признать наличие и “доминантной” цикличности [2]. Видимо, в этом сомневаться не приходится.
В статьях Г.В. Казеева “Учение о половом цикле” приводится этологический подход к ПЦ в скотоводстве, в котором вместо полового возбуждения или общей реакции выделены поведенческие реакции: “поисковая и ритуал ухаживания “ при этом половая охота также относится к поведенческим реакциям и критикуется (с позиций автора имеется “вечная ошибка” в официальных нормативных документах-инструкциях МСХ РФ, рекомендациях и др.) утверждение, что рефлекс неподвижности и половая охота являются равнозначными, тождественными состояниями. Игнорируя работу академика В.К.Милованова (1962), детально описавшего локомоторные реакции животных при осуществлении половой функции, автор считает, что его данные (введение ритуала ухаживания феномен половой охоты, который ничего не дает производственникам, так как во всех странах применяется принцип “до полудня – после полудня”, который обеспечивает оптимальную оплодотворяемость: «см. главу 13 [1] ») дополняют и расширяют учения о ПЦ А.П. Студенцова и его последователя В.С.Шипилова (1986) и сожалеет, что в современных условиях, в мировом масштабе на производстве нет быков-пробников и самым распространенным методом выявления сроков осеменения коров является визуальный, отрицая который учение А.П.Студенцова в какой-то мере теряет притягательную силу для специалистов. Рекомендация проводить исследование цервикальной слизи на производстве маловыполнима, так как на это у специалистов не бывает времени и примерно в 47% случаев слизь не удается получить для исследования. Специалист скорее применит HEAT-DETECTOR, чем будет исследовать слизь [27].
В учебнике, изданном под ред. академика В.Я.Никитина и проф. М.Г.Миролюбова в 2000 г., повторяется определение данное А.П.Студенцовым, при этом учебник отстает от современного уровня на 30-40 лет, насыщен многочисленными неточностями и неясностями, противоречивыми понятиями и излишними рассуждениями, например, ”течка характеризуется ярко выраженными пролиферативными процессами, ”(тогда, где голокриновая секреция эпителиоцитов шейки матки?); без объяснения утверждается, что “различают течку первой, второй и третьей степени”; “когда яйцевая клетка почему-либо не участвует в оплодотворении или зигота не прививается”(?); “взаимосвязь течки, полового возбуждения, охоты и овуляции во времени благоприятствует оплодотворению” (но как?); “фолликулы, подвергающиеся атрезии, выделяют фолликулин” (а растущий, доминантный не известно -?; при этом нигде не упоминается Е2); “фолликулярный гормон действует только на проводящие пути полового аппарата”(?); а дальше опровергают себя, указывая, что “накопление в организме фолликулина вызывает реакцию нервной системы, проявляющуюся половым возбуждением и охотой”; опровергая эстральный цикл авторы пишут – “при исследовании яичника удается проследить”, “различить фазы фолликуло – и лютеогенеза”(?), и т.д. [28].
В учебных пособиях (1995, 1997 гг.) группы авторов [21,22], во главе с В.В.Ельчаниновым, критикуется “нервизм” И.П.Павлова, А.П.Студенцова за термины “рефлекс охоты, течки и овуляции”, за стадии возбуждения, торможения и уравновешивания”, работы А.А.Ухтомского и В.К.Милованова за применение терминов “половая доминанта”, “доминанта беременности”, “ лактационная доминанта”, опровергается учение М.М.Сеченова фразой, что “в современных учебниках, благодаря инерционности мышления, кора головного мозга по-прежнему представлена в качестве центрального регуляторного органа репродуктивной функции”, что “половое поведение животных нельзя втиснуть в рамки понятия “рефлекс”, что “пища, свет и самец поставлены в один ряд как раздражители коры головного мозга”. В противовес провозглашается “полуавтономность регуляторной триады – гипоталамус – гипофиз – гонады”. Обсуждая вопросы эстрального цикла (подчеркиваем “не менструального”!) авторы насыщают свои руководства терминами из медицины и зоопсихологии, не принятые в животноводстве, например: “оргазм”, ”эмоциональный”, ”препубертат”, ”пубертат”, ”дериваты”, “триместр внутриутробного развития”, “адреналова железа”, “плодового организма”(?), “гипоталамический гонадостат”(?), “нимфоманизм”, “самцового типа” и т.д. Непонятно, зачем вводить в заблуждение специалистов этими словами, ратуя за “грамотный подход” к объяснению нарушений функции размножения, при этом совершенно неграмотно объясняется гипофункция яичников при обнаружении в них ЖТ, Ф-3, Ф-2, а при обнаружении мелких Ф-1 – оценивается как “глубокая форма гипофункции яичников”(может быть она менее глубокая?), т.к. рекомендуется овариоскопия и послеубойный осмотр яичников (применяют ГСЖК и PGF2альфа). Достаточно прочитать заголовок этой главы 4.1 “Задержка пубертата. Гипофункция яичников инфантильного и препубертатного типа, методы гормональной стимуляции полового созревания”(?), чтобы понять насколько далеки авторы от реального животноводства [22].
Академик Л.К.Эрнст и А.Н.Варнавский (2002) определяют ПЦ самок как “способность неоплодотворенной самки приходить в состояние половой охоты (либидо) через определенные промежутки времени”, ПЦ самок можно разделить на 4 стадии: проэструс, эструс, “метаэструс” и диэструс; по нервным процессам ПЦ делится на 3 стадии: возбуждение, торможение и уравновешивание, т.е. здесь смешивается два разных определения, и лишь дальше заключается: “ПЦ, таким образом”, состоит из чередования относительно длительной фазы ЖТ (вырабатывается прогестерон) и непродолжительной фолликулярной фазы (зрелый фолликул вырабатывает эстрогены). Необоснованно показана концентрация гормонов в минус первой степени, и подразделение желез в эндометрии (“матке”) на четыре типа, введение нового термина – “алибидии”, утверждение, что “первая после отела полноценная половая охота наиболее плодотворная”. И, наконец, обоснование полноценности - “Полноценность охоты надо дифференцировать по продолжительности – она должна быть не менее 17 дней “, (что в корне неверно) и заключение, что “в России 27% коров иммунно бесплодны”(?)/ [29,с.262].
Допускаются мистические и неясные выражения: “при наличии в крови достаточного количества П окситоцин теряет свою силу”(?), “половая и материнская доминанты вступают в конфликт, в борьбу”; ”нужно вмешательство в борьбу половой и лактационной доминант, при которой половая система остается в анемичном, дистрофическом (?) состоянии”; “гипоталамус подразделяется на низшие и высшие центры”(?); “нейросекрет супраоптических и паравентрикулярных ядер достигает аденогипофиза(?)”, а через десяток страниц правильно утверждается, что он опускается по аксонам в нейрогипофиз; ”за счет маточного продукта, подобного эстрадиолу (?); обеспечивается синхронная функция гипоталамуса, гипофиза, яичников и матки(?); рассматривается роль различных нейромедиаторов (которые неправильно называются “хемомедиаторами”) и признается саморегулирование по принципу обратной связи, т.е. теория “плюс-минус взаимодействия” М.М.Завадовского, но затем он критикуется за то, что “пытался вывести общий закон взаимодействия эндокринных желез” и т.д.[29].
Приведенный обзор представлений разных авторов о ПЦ показывает насколько разный подход к этому вопросу имеется у авторов разных школ в России, вплоть до эклектического смешивания и противопоставления аргументов разных ученых. Апогеем такого подхода является работа А.Г.Нежданова (2003), в которой хаотически смешиваются разные понятия и противопоставляются друг другу: автор использует, а затем игнорирует работы М.М.Завадовского (1929-1933), о “плюс-минус взаимодействии”, взятые за основу в теории функциональных систем П.К.Анохина (1930-1934); вынужден признать циклические изменения, связанные с яичниковым или овариальным циклом; принижает роль рефлекторной теории, опираясь на утверждения К.В.Судакова, абсолютизирующие влияние гипоталамических структур на кору головного мозга и устраняющие примат внешней стимуляции, и, наконец, по примеру партийного руководства советского периода, бросает угрозу, что если найдутся инакомыслящие “в этой области, то их старания равносильны чернительству мертвого льва”! [30]. Таким образом, по-старинке разные школы ученых, вместо разработки современных представлений о репродуктивной функции, по-прежнему противопоставляются друг другу, как следствие августовской сессии ВАСХНИЛ (1948) и “Павловской” сессии АМН СССР (1950).
Многочисленные клинические и экспериментальные исследования дали возможность существенно расширить представления об основных закономерностях процессов роста фолликула, овуляции и развития ЖТ, охарактеризовать особенности гипофизарной гонадотропной и гипоталамической регуляции этих процессов. Полученные данные в совокупности с результатами изучения секреции половых (гонадальных) и гонадотропных гормонов гипофиза послужили основой для создания стройной концепции о системе взаимосвязей, обеспечивающих половую цикличность репродуктивной системы самок животных (рис. 12).
Цикличность деятельности яичников зависит от образования двух временных эндокринных структур – зрелого граафового (доминантного) фолликула и образующегося после овуляции желтого тела. Длительность цикла у животных и все его параметры сильно зависят от продолжительности жизни фолликула и желтого тела, обуславливающих влияние гормонов яичника на гипоталамус, аденогипофиз и матку через механизмы обратной связи. Только зрелый фолликул секретирует достаточное количество эстрогенов, которые могут стимулировать гипоталамо-гипофизарную систему к выделению ЛГ в количестве, достаточном для того, чтобы вызвать разрыв фолликула и образование желтого тела. У каждого вида продолжительность лютеальной фазы сравнительно постоянна. Поскольку гормоны ЖТ подавляют секрецию гипофизом гонадотропных гормонов, желтое тело регулирует также развитие фолликулов.
Ведущую роль в проявлении функции размножения выполняет центральная нервная система, регулирующая секрецию гормонов и опосредующая их обратное действие. Внешние раздражители (триада А.П. Студенцова: корм, свет, самец) поступают через анализаторы (зрительный, обонятельный, слуховой, вкусовой, осязательный), внутренние раздражители – гормоны, нейромедиаторы, сигнализируют о готовности животного к размножению. Головной мозг суммирует полученные раздражения и направляет нервные импульсы гипоталамусу, где они преобразуются в гуморальные, способствующие выделению гонадотропных гормонов гипофиза. Возбужденный гипоталамус посредством ретикулярной формации индуцирует (настраивает) кору головного мозга, вызывая в ней сексуальную настроенность в поведении самки или самца, т. е. формирует половую доминанту – временный господствующий очаг возбуждения в центральной нервной системе. При этом многие функции организма мобилизуются на осуществление воспроизведения.
В регулировании репродуктивной функции прослеживается пять уровней регуляции по иерархическому типу: нижележащие структуры регулируются вышележащими, которые, в свою очередь, реагируют на изменения в нижележащих уровнях по принципу обратной связи. При этом сигналы, поступающие от нижележащих структур, корректируют деятельность вышележащих. (см. Рис.12).
Специфические нейросекреторные факторы (гонадолиберины – ГЛ и статины), вырабатывающиеся в гипоталамусе поступают по воротной (портальной) кровеносной системе в переднюю долю гипофиза и стимулируют выделение гонадотропных гормонов (тоническая секреция ФСГ и ЛГ) аденогипофизом. Гонадотрофы увеличивают выделение ФСГ и ЛГ, которые гуморальным путем попадают в яичники и стимулируют развитие вторичного фолликула в преовуляторный (доминантный) или третичный граафов фолликул.
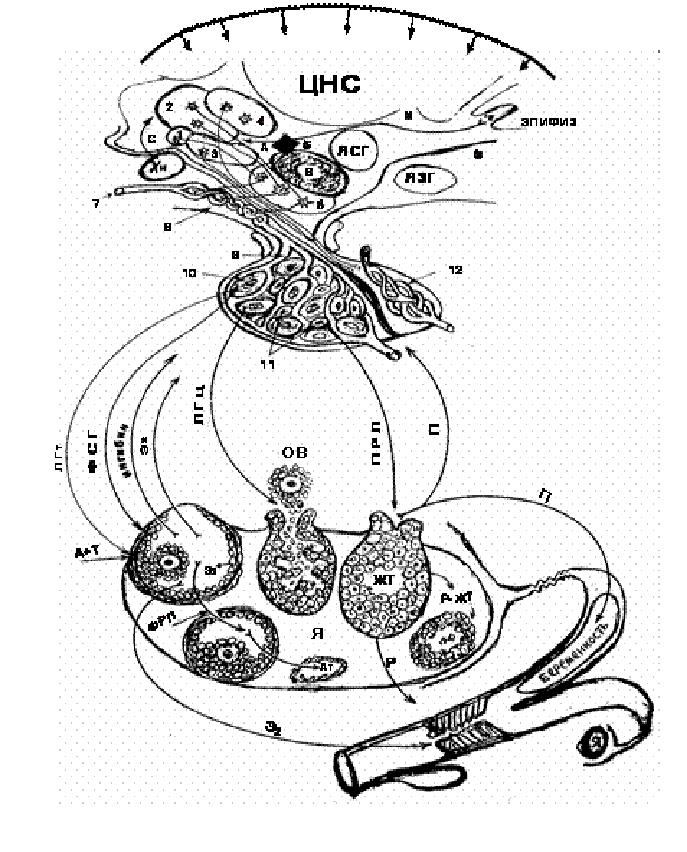
Рис. 12.Нейрогуморальная регуляция половой функции: А – ядра переднего гипоталамуса: 1 – супрахиазматическое, 2 – преоптическое, 3 – супраоптическое, 4 – паравентрикулярное; Б – ядра среднего гипоталамуса: 5 – вентромедиальное, 6 – аркуатное; ЯСГ – другие ядра среднего гипоталамуса; В – ЯЗГ – ядра заднего гипоталамуса (комплекс мамиллярных ядер); 7 – верхняя гипофизарная артерия; 8 – медиальная эминенция с первичной капиллярной сетью и капиллярными петлями; 9 – портальные сосуды гипофиза (аденогипофиза); 10 – гонадотрофы; 11 – лактотрофы;12 – портальные сосуды нейрогипофиза; А – Б – полость третьего мозгового желудочка; Хи – хиазма зрительных нервов; М – мелатонин – гормон эпифиза; Е2 или Э2 - эстрадиол; С – серотонин; Р – релаксин.
На первичные фолликулы гонадотропины не действуют. В преовуляторном фолликуле синтезируются эстрогены, главным образом, 17-бета- эстрадиол (Е2) и небольшое количество эстрона, однако эстрон и эстриол образуются преимущественно в печени из эстрадиола, а также в других тканях из андрогенов, главным образом из андростендиона. (рис.11) ЛГ взаимодействует с рецепторами на клетках теки, стимулируя выработку андрогенов и небольших количеств эстрадиола. ФСГ активирует систему ферментов, осуществляющих ароматизацию в гранулезных клетках, и превращает большую часть андрогенов (А+Т) в эстрогены. Таким образом, синтез эстрогенов в фолликулах регулируется ФСГ, действие которого опосредуется циклическим аденозин-монофосфатом (цАМФ). Местный эффект стероидных гормонов реализуется путем пассивной диффузии их в любые клетки организма, но задерживаются они лишь в клетках гормон-чувствительных тканей, где эстрогены образуют комплексы с цитозольными рецепторами с последующей транслокацией в ядро клетки и воздействием на генетический аппарат. Образующиеся эстрогены гуморальным путем подавляют выделение ФСГ из аденогипофиза по принципу обратной связи и стимулируют выделение циклического ЛГ (ЛГц). Концентрация Е2 в полости развивающегося фолликула в 1000 раз превышает его концентрацию в кровотоке, что локально создает условия для развития доминантного фолликула. Пик увеличения количества эстрогенов в крови наблюдается между 48-м и 24-м часами до начала охоты.
Созревающий доминантный фолликул продуцирует возрастающие количества ингибина и фолликулярного регулирующего протеина (ФРП). Ингибин, воздействуя на гипофиз, способствует понижению секреции ФСГ и, таким образом, блокирует появление новых крупных фолликулов, а ФРП из преовуляторных фолликулов угнетает синтез эстрадиола во второстепенных фолликулах (Ф2), что приводит к их атрезии (Ат), т. е. обратному развитию без овуляции. Повышение содержания Е2 активизирует гипоталамус, который стимулирует выделение из аденогипофиза большого количества овуляторного – циклического ЛГ (ЛГц). В ответ на резкое возрастание количества ЛГ в плазме усиливается рост клеток гранулезы фолликула, начинается секреция прогестерона (П), PGF2альфа и PGЕ2, и высвобождение коллагенолитического фермента, истончающего стенку фолликула, в результате образуется стигма (бессосудистая область), где происходит овуляция. Важную роль играет и другой протеолитический фермент – активатор плазминогена, который способствует отделению ооцита от яйценосного бугорка.
Выброс циклического ЛГ у коров продолжается 6–12 часов, а период его полужизни в циркулирующей крови составляет 35 минут. Величина преовуляторного пика ЛГ в сыворотке крови коров молочных пород достигает в среднем 12,9 нг/мл (5,0–19,5 нг/мл), у телок – 25,9 нг/мл (19–35 нг/мл). Базальный уровень ЛГ составляет 1,0–2,0 нг/мл (4,6–8,5 нг/мл). Овуляция наступает через 10–12 часов после окончания охоты или через 25–30 часов после предовуляторного выброса ЛГ, когда фолликул достигает полного развития и размеров 16–18 мм в диаметре. М. И. Прокофьев (1979) выявил прямую зависимость между оплодотворяемостью коров и уровнем ЛГ в их крови в предовуляторный период [16].
Результаты анализа гонадотропной функции гипофиза, секреции эстрогенов и морфологических исследований фолликулярной системы яичников позволяют выявить несколько физиологических ритмов в функционировании гипоталамо – гипофизарного комплекса и яичников крупного рогатого скота в течение полового цикла.
У коров концентрация прогестерона в крови быстро падает в период между 72-м и 48-м часами, предшествующими охоте. Взятие крови каждые 4 часа в течение 24 часов до и после начала охоты позволило обнаружить подъем уровня прогестерона в крови коров за 16 часов до начала охоты. С 4-го до 12-го дня ПЦ уровень прогестерона в плазме крови коров быстро повышается, достигая 4-5 нг/мл и остается на этом уровне до 16-го дня ПЦ.
Уровень эстрогенов в крови коров начинает повышаться за 4 дня и достигает пика за 24 часа до начала охоты, а затем снижается в течение 48 часов до базального уровня. Таким образом, у коров, как и у овец, повышенная секреция эстрогенов предшествует предовуляторному выбросу циклического ЛГ, что дает основание считать их «физиологическим пускателем» выделения ЛГ (Рис.13).
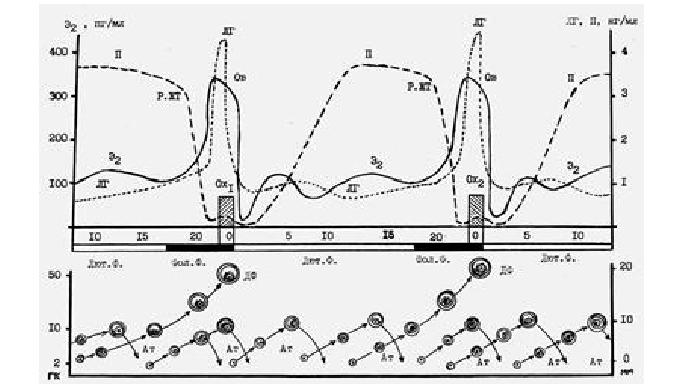
Рис. 13. Динамика изменения секреции гормонов яичника, гипофиза и роста фолликулов в течение полового цикла коров молочных пород:Э2 (Е2),– эстрадиол, пг/мл; П, ЛГ = концентрация прогестерона и лютеинизирующего гормона, нг/мл; Фол. – Ф. – фолликулярная фаза ПЦ;Лют. Ф. – лютеальная фаза ПЦ; Р-ЖТ – регрессия ЖТ; Ов – овуляция; Ох1 – охота первого ПЦ; Ох2 – охота второго ПЦ; Внизу: слева – число гранулезных клеток (ГК) в фолликуле х 106; справа – диаметр фолликулов, мм; Ат – атрезия фолликулов; ДФ – доминантный фолликул.
Второй подъем уровня эстрогенов установлен в период начала ПЦ с 1-го до 5-го дня, и даже 6-го–8-го дней, до того как ЖТ начинает секретировать большое количество прогестерона.
Третий пик концентрации эстрогенов в плазме крови коров найден на 13–16-й день ПЦ перед регрессией ЖТ, хотя уровень Е2 оставался относительно низким. Именно в эти периоды ПЦ, на 5–6-й, 12–14-й и 18–19-й дни наблюдается заметное увеличение числа крупных фолликулов в яичниках коров.
На месте овулировавшего фолликула образуется ЖТ, выделяемый им прогестерон, по принципу обратной связи угнетает выделение ГЛ, поэтому аденогипофиз выделяет только тонический уровень ЛГ. Овуляция новых фолликулов не происходит. Под влиянием прогестерона эндометрий подготавливается к приему и прикреплению зиготы и обеспечивает нормальное протекание беременности. При отсутствии в матке зародыша эндометрий выделяет лютеолитический фактор – простагландин (PGF2альфа), который по противоточному механизму поступает в яичник и вызывает регрессию ЖТ. Выделение прогестерона резко уменьшается, на фоне наличия в яичниках новых растущих фолликулов. Рост их начинается сразу же после овуляции вследствие уменьшения уровня ингибина, фолликулярного регулирующего протеина, эстрадиола и последующего периодического увеличения выделения ФСГ. У крупного рогатого скота первая после овуляции волна роста фолликулов заканчивается атрезией вначале мелких, а затем и крупных фолликулов. Атрезия, в свою очередь, дает начало новой волне роста фолликулов на 5–6-й и 12–14-й дни ПЦ. В результате последней волны на 18–19-й дни появляется доминантный овуляторный фолликул и происходит овуляция. Следовательно, завершается предыдущий и начинается новый ПЦ, а в случае успешного осеменения животного развивается беременность. У беременных животных лютеолитический эффект эндометрия устраняется присутствием эмбриона и ЖТ беременности сохраняется на протяжении всего периода стельности. Прогестерон активирует нервные центры, связанные с материнством, и формируется материнская доминанта. В результате поведение самки меняется: течка и охота прекращаются, самка не подпускает самца для спаривания, в матке начинается подготовка к прикреплению зародыша, эмбриональному и плодному периоду его развития.
В течение цикла воспроизводства у крупного рогатого скота наблюдается несколько половых циклов до возникновения беременности. На количество ПЦ в значительной мере влияют условия внешней среды: температура, кормление, содержание. Разнообразное и полноценное кормление самок создает условия для обеспечения нормальных гормональных параметров крови, необходимых для функционирования всей репродуктивной системы. Внешние и внутренние факторы (находятся под коррелирующим влиянием ЦНС, включающей кору головного мозга), обеспечивают безусловно-рефлекторную функцию размножения, при этом важнейшая роль в цепи нейрогуморальных регуляторных механизмов, принадлежит сложнейшему гормональному ансамблю в гипоталамо-гипофизарно-овариальной системе с ее прямыми и обратными связями.
Литература
1. Середин В.А. “Биотехнология воспроизводства в скотоводстве: учебное пособие. - Нальчик: ИЦ ”Эль-Фа”, 2004. - 472 с., ил.
2. Шипилов В.С. Физиологические основы профилактики бесплодия коров.- М.: Колос, 1977. - 336 с., ил.
3. Завадовский М.М. Теория и практика гормонального метода стимуляции многоплодия сельскохозяйственных животных.- М.: Сельхозиздат, 1963.- 677с., ил.
4. Биохимия: Учебник / Под ред. Е.С.Северина.-2-е изд. – М.: ГЭОТАР-МЕД, 2004.- 784 с., ил.
5. Руководство по эндокринологии / Под ред. Б.В.Алешина, С.Г.Генеса, В.Г.Вогралика. - М.: Медицина, 1973. - 512 с.
6. Бородулин Ф.Р. С.П.Боткин и неврогенная теория медицины. – 2-е изд. – М.: Медгиз, 1953. – 184 с.
7. Павлов И.П. Лекции по физиологии 1912-1913 гг. / Зап. и сист. П.С.Купановым; Под общ. Ред. И.П.Разенкова. – М.: Изд-во АМН, 1952. - 331 с.
8. Сеченов И.М. Рефлексы головного мозга.- М.: Изд-во АМН, 1952.- 231 с.
9. Коренной Н.И., Дьяченко П. Важный фактор определения времени осеменения коров // Молочное и мясное скотоводство. -1974. - №4.- С. 28, 29.
10. Павлов И.П. Двадцатипятилетний опыт объективного изучения высшей нервной деятельности (поведения) животных. Условные рефлексы.- М-Л.: Биомедгиз, 1938.- 771 с., ил.
11. Милованов В.К. Биология воспроизводства и искусственное осеменение животных. - М.: Сельхозиздат, 1962. - 696 с.
12. Студенцов А.П. Диагностика беременности и бесплодия с.-х. животных.-М.: Сельхозгиз, 1949,- 1950. - 102 и 135 с.
13. Быков К.М. Учение И.П.Павлова и современное естествознание.- М.: Медгиз, 1952. - 36 с.
14. Студенцов А.П. Ветеринарное акушерство и гинекология. -М.: Сельхозгиз, 1953. - 502 с.
15. Руководство по эндокринной гинекологии / Под ред. Е.М.Вихляевой. - М.: Мед.информиздат, 1997.- 768 с., ил.
16. Прокофьев М.И. Регуляция размножения сельскохозяйственных животных.- Л.: Наука, 1983.-264 с.
17. Прохорова В. Ультразвуковая диагностика // Ветеринарная газета.-Сентябрь 1999.- №17(165).- С. 10-11 с.
18. Гормональная регуляция размножения у млекопитающих: Пер. с англ. / Под ред. К.Остина и Р.Шорта.- М.: Мир, 1987.- 305 с., ил.
19. Иванов И.И. Избранные труды. - М.: Колос,1970.- 320 с.
20. Гормональная регуляция размножения у млекопитающих: Пер. с англ. / Под ред. К.Остина и Р.Шорта.- М.: Мир, 1987.- 305 с., ил.
21. Ельчанинов В.В., Белоножкин В.П., Насибов Ш.Н. Проблемы физиологии и патологии репродуктивной функции коров / Ч.I. Быково, Подольск. р-он, Московск. обл.: Щербинская типогр., 1997.- 291 с.
22. Ельчанинов В.В., Тяпугин Е.А. Формирование репродуктивной функции у самцов и самок с.-х. животных в онтогенезе. - М.: АОЗГ “Зоосалон”, 1995.- 120 с.
23. Сысоев А.А. Физиология размножения сельскохозяйственных животных. - М.: Колос, 1978.- 360 с., ил.
24. Сергиенко А.И. Профилактика бесплодия крупного рогатого скота.- М.: Колос, 1984.- 188 с., ил.
25. Валюшкин К.Д., Медведев Г.Ф. Акушерство, гинекология и биотехника размножения животных.- Минск: “Ураджай”, 1997.- 718 с.
26. Солсбери Г.У., Ван-Демарк Н.Л. Теория и практика искусственного осеменения коров в США / Под ред. и предисл. Акад. ВАСХНИЛ В.К.Милованова. - М.: “Колос”, 1966.- 527 с.
27. Казеев Г. Учения о половом цикле – теоретическое обоснование организации искусственного осеменения коров// Животновод. – 2002. - №7-8. – С.10-12; №9. – С.8-10; №10. – С.10-12.
28. Ветеринарное акушерство, гинекология и биотехника размножения / А.П. Студенцов, В.С.Шипилов, В.Я.Никитин и др.- М.: Колос, 2000.- 495 с.
29. Эрнст Л.К., Варнавский А.Н. Репродукция животных. -М.: Биотех, 2002.- 364 с.
30. Нежданов А.Г.Современное представление о половом цикле самок животных // Ветеринария.- 2003. - №11.- С.32-36.
Алексей Киселев - организация пункта искусственного осеменения
drakon060976@mail.ru





