
ОБРАЗОВАНИЕ МОЛОКА В МОЛОЧНЫХ ЖЕЛЕЗАХ
И ПЕРИОД ЛАКТАЦИИ
КРИВАЯ ЛАКТАЦИИ
НАЧАЛО ЛАКТАЦИИ
Во время отела взрослой коровы или телки, в тканях молочных желез происходят существенные физиологические изменения. Еще до отела ткани, вырабатывающие молоко, уже развиты и готовы к производству молока, но они как бы отключены до тех пор, пока не получат соответствующие гормональные сигналы при родах. Эти сигналы поступают как от самой коровы, так и от исзчезновения гормонов, вырабатывающихся плацентой. Все вырабатывающие молоко ткани имеются в наличии и готовы к работе до отела, но не могут функционировать, пока не "включены" при отеле.
• Передняя доля гипофиза коровы выделяет всплеск пролактина, и в это же время наблюдается возрастание уровня содержания гормона роста в крови. Яичники коровы увеличивают выделение эстрогена и прекращают остаточное выделение. • Отделение плаценты устраняет источник плацентарного лактогена, служащего для стимуляции развития молочных желез в период беременности, а также важный источник прогестерона, поддерживавший беременность на поздних стадиях.
Первоначальная стимуляция, вызывающая эти изменения, исходит по-видимому от плода, повышение выделения гормона коры надпочечников которого, видимо, инициирует отделение плаценты. Ни один из упомянутых сигналов в отдельности не вызывает полное начало лактации; для этого необходимо их совместное воздействие.
По мере того, как изменения происходят в секреторной способности
ткани молочных желез, начинается также мобилизация ферментов, перерабатывающих сырье в различные компоненты молока: белки, углеводы, жиры и другие составляющие.
Для начала лактации при отеле необходимо сложное взаимодействие гормональных изменений.
Первоначальный импульс исходит от плода.
Прогестерон подавляет образования фермента синтетазы лактозы, необходимого для выработки лактозы. Этот фермент состоит из двух компонент. Одной из них является а-лактальбумин, который образуется при снижении уровня прогестерона. Как только фермент синтетазы лактозы полностью сформирован, становится возможным синтез лактозы в альвеолах. Он же вызывает втягивание в альвеолы воды из крови, обеспечивая тем самым наличие жидкого носителя для других компонент молока.
КРИВАЯ ЛАКТАЦИИ
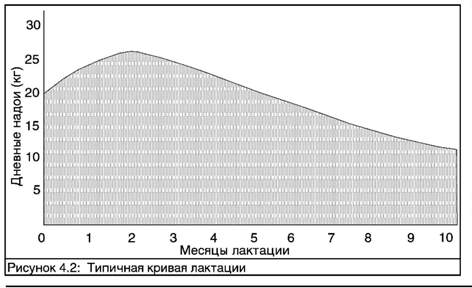
Производство молока нарастает в первые три-шесть недель лактации, а затем постепенно спадает. Обычно коров намеренно проводят через период сухостоя в течение двух месяцев перед следующим отелом. Этот период отдыха необходим для достижения максимальной молочной продуктивности в следующей лактации. Реальное количество произведенного молока и форма кривой лактации зависит частично от сезона отела, географического региона и нескольких факторов содержания, включающих питание и частоту доения.
Производство молока возрастает в течение нескольких первых недель лактации, а затем постепенно спадает.
Поскольку форма кривой сравнительно постоянна, молочная продуктивность в начальной стадии лактации может быть использована для вычисления полной продуктивности за всю лактацию. Таблица 4.1 дает ориентир для оценки полного надоя за лактацию (305 дней).
ОТДАЧА МОЛОКА
В природе молоко выходит, когда теленок сосет. В молочном стаде корову необходимо тренировать, чтобы она реагировала на сигналы в неестественных для нее условиях.
Миоэпителиальные клетки (гладкие мышцы) действуют под контролем гормонов. При стимуляции они сокращаются, выдавливая молоко в систему протоков вымени. Этот гормональный сигнал инициируется комбинацией нервной и гормональной реакций. Коровы реагирует на стимуляторы нервной системы (ощущения), сообщающие, что теленок готов сосать или что доение начинается. Такими сигналами могут быть визуальные - вид теленка; звуковые - звуки, производимые теленком, или шум, ассоциируемый с доением; или тактильные сигналы -ощущение теленка касающегося вымени или обмыв вымени перед дойкой. В ответ на такие нервные стимуляторы
Таблица 4.1: Ориентир для оценки полного надоя за лактацию (305 дней).*
|
Месяц лактации |
Число дней в дойке |
Факторы для предсказания полногонадоя за лактацию |
|
|
|
|
Первотелки в первой лактации |
Коровы во второй лактации и старше |
|
1 |
16 |
0.348% |
0.371% |
|
2 |
46 |
0.409% |
0.421% |
|
3 |
77 |
0.397% |
0.400% |
|
4 |
107 |
0.381% |
0.376% |
|
5 |
138 |
0.362% |
0.350% |
|
6 |
168 |
0.344% |
0.326% |
|
7 |
199 |
0.323% |
0.299% |
|
8 |
229 |
0.301% |
0.276% |
|
9 |
260 |
0.277% |
0.248% |
|
10 |
290 |
0.249% |
0.211% |
* Выберите месяц и среднюю точку и разделите надой этого дня на соответствующий процентный показатель. Например, взрослая корова, давшая 25 кг в 77-й день должна дать приблизительно 25/0,400x100=6250 кг за 305 дней.
задняя доля гипофиза выделяет гормон окситоцин. Окситоцин переносится с кровотоком и связывается с миоэпителиальными клетками, которые сокращаясь вызывают выделение молока.
|
Нервные и гормональные реакции контролируют выделение молока. |
Такие нейроэндокринные (нервно-гормональные) рефлексы могут являться хорошей защитной мерой в процессе эволюции - молоко не "тратится", когда теленок не готов сосать. Отрицательные нервные стимуляторы, такие как незнакомые шумы или боль, могут подавлять выделение молока. Этот эффект вызывается другим гормоном адреналином. Таким образом, дойка должна происходить в спокойной, знакомой животному обстановке. Громкие звуки, крики или удары могут уменьшить выделение и, соответственно, надой молока.
Европейские породы коров и породы типа зебу довольно заметно различаются по реакции выделения
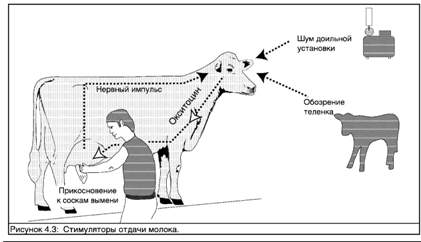
молока. Тогда как основные европейские молочные породы, например голштинская и джерсийская, хорошо реагируют на искусственные стимуляторы, предшествующие машинному или ручному доению, коровам зебу для начала выделения молока может требоваться визуальный или действительный контакт со своим собственным теленком. Это, очевидно, делает их менее адаптируемыми к доению в условиях доильного зала при машинном доении. В некоторых системах молочного производства теленку позволяют сосать один сосок, в то время как остальные три четверти вымени доятся вручную. Среди других "трюков" инициации выделения молока у коров зебу встречается привязывание теленка у головы коровы, в то время как она доится с помощью машинного аппарата.
Выделение молока затруднено, если корова нервничает или напугана;
Спокойная, тихая обстановка необходима для получения максимального выделения молока.
ПРЕКРАЩЕНИЕ ЛАКТАЦИИ ИЛИ ЗАПУСК КОРОВЫ
Гормон окситоцин важен не только для выделения молока, но и для поддержания лактации. Производство молока продолжается до тех пор, пока оно регулярно удаляется из молочных желез. Поэтому корова, переодически испытывающая стимуляцию с выделению молока, будет продолжать его производить, хотя надои с продолжением лактации в конце концов упадут.
Если молоко больше не удаляется регулярно из вымени, то секреторные ткани претерпевают инволюцию (обратное развитие). Это происходит, когда теленка отлучают от матери или частоту доения намеренно снижают до одного раза в день, а затем прекращают доение.
Если молоко больше не удаляется из вымени, то секреторные ткани прекращают производство молока.
Часть произведенного молока при инволюции поглощается, и клеточные структуры проходят процесс остановки, в котором их метаболическая активность, число секреторных клеток и размер каждой доли уменьшаются. Постепенно альвеоли распадаются, клетки проникают в железы и удаляют остатки из эпителиальных клеток, оставляя миоэпителиальные клетки нетронутыми. В результате инволюции система протоков остается, но альвеоли сжимаются. У беременных коров процесс инволюции подавляется необходимостью развития молочных желез для их подготовки к производству молока в новой лактации.
КАКОЙ ЭФФЕКТ ОКАЗЫВАЕТ ПЕРИОД СУХОСТОЯ НА ПОСЛЕДУЮЩУЮ ЛАКТАЦИЮ?
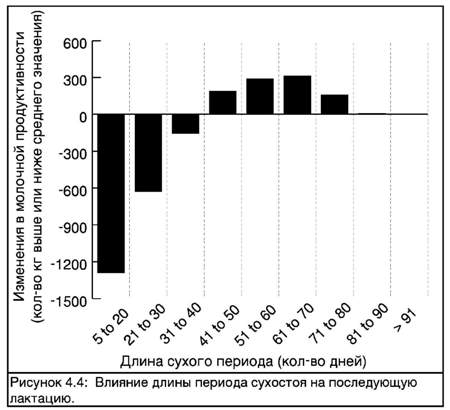
Исследования показали, что когда период сухостоя продолжается менее 40 дней, это оказывает негативное влияние на молочную продуктивность в последующей лактации. Однако период сухостоя продолжительностью более 80 дней может также немного снизить продуктивность. Рисунок 4.4 демонстрирует влияние изменения продолжительности периода сухостоя на молочную продуктивность.
ДРУГИЕ ФАКТОРЫ, ВЛИЯЮЩИЕ НА ПРОДУКТИВНОСТЬ ЗА ВРЕМЯ ЛАКТАЦИИ
Порода
Среди обычных молочных пород, разводимых в Соединенных Штатах, голштинская порода лидирует по общему объему надоев и общему выходу
всех основных компонент молока. Молочная продуктивность и состав молока могут значительно меняться у различных особей внутри одной породы. Этот факт служит базисом для селекционного разведения и более подробно обсуждается в Техническом Руководстве "Воспроизводство и Генетическая Селекция".
Порядковый номер лактации
Производство молока возрастает с порядковым номером лактации и достигает максимума на четвертую или пятую лактацию. Это происходит в результате развития и возрастания размера вымени, а также увеличения размера тела животного по сравнению со временем первой лактации. Размер тела влияет на способность животного потреблять достаточное количество питательных веществ. Надой в певой лактации для животного, отелившегося в возрасте двух лет, нужно умножить приблизительно на 1,3, чтобы получить "взрослый эквивалент" надоя, или другими словами - что можно ожидать от коровы, когда она полностью повзрослеет. Поправочный коэффициент меняется в зависимости от породы коров, сезона отела и региона.
Месяц отела
Было установлено, что в типичных условиях фермы Среднего Запада Соединенных Штатов коровы, телящиеся в конце зимы дают приблизительно на 8% больше молока, чем коровы, телящиеся летом. Сходные сезонные изменения наблюдаются в других регионах мира. Они определяются взаимовлиянием доступности кормов, фотопериодом (длиной светового дня) и температурой окружающей среды.
Факторы содержания
Кормление и доступ к воде
Метаболизм тканей молочных желез очень высок, что позволяет им вырабатывать огромное количество питательных веществ, содержащихся в молоке. Любое снижение количества сырья, попадающего в вымя, приводит к падению молочной продуктивности. Наиболее заметное влияние оказывает нехватка воды, поскольку организм коровы не может запасать воду. Ограничение доступа к воде или подача ее в недостаточном количестве в течение нескольких часов приведет к быстрому падению молочной продуктивности. На ранней стадии лактации (приблизительно до 20й недели) корова может мобилизовывать запасы жира и белков в организме для компенсации нехватки питательных веществ в рационе. Потребности коров в питании на разных стадиях лактациях обсуждаются в Техническом Руководстве Пищеварение и Кормление (Глава 6).
Недостаток воды, даже в течение всего нескольких часов, вызовет резкое падение молочной продуктивности.
Гормон роста
Уже более двадцати лет является известным тот факт, что существует взаимосвязь между молочной продуктивностью коров и уровнем содержания гормона роста в крови. Поэтому животные с большим генетическим потенциалом к производству молока обычно имеют более высокое содержание этого гормона в крови. Гормон роста выполняет множество функций в организме, и еще не до конца выяснено, которая из них стимулирует лактацию. Многие органы обладают рецепторами гормона роста и некоторые из них, как например печень, связывают гормон роста и в свою очередь вырабатывают вторичный гормон (IGF-I). Поскольку метаболический эффект, оказываемый гормоном роста, хорошо описан, считается, что он вызывает такое перераспределение питательных веществ в организме коровы, которое благориятствует их использованию для производства молока. Не наблюдается связи гормона роста секреторными клетками и какого-либо прямого эффекта, оказываемого им на секрецию молока. Одно из исследований выявило возрастания содержания а-лактальбумина, хотя общее содержание белка в молоке существенно не изменилось. Изменение уровня а-лактальбумина способствует возрастанию выделения лактозы и тем самым связано с увеличением движения воды, повышая таким образом производство молока.
СЕКРЕЦИЯ НА КЛЕТОЧНОМ УРОВНЕ
В синтезе молока участвует весь организм коровы. Лактация зависит не только от процесса воспроизводства, но и от правильного питания коровы, которое либо должно обеспечивать достаточное количество питательных веществ, или сырья, для синтеза молока, либо делать возможной мобилизацию этих веществ из других источников в организме. Информация о том, как питательные вещества попадают в кровь и доставляются в молочные железы, содержится в Техническом Руководстве "Пищеварение и Кормление". Нижеследующий раздел освещает только метаболизм компонентов молока в секреторных клетках тканей молочных желез.
СИНТЕЗ УГЛЕВОДОВ МОЛОКА
Углеводы, содержащиеся в рационе коровы, в результате ферментации в желудке коровы превращаются в летучие жирные кислоты (ЛЖК), включающие уксусную, пропионовую и масляную кислоты (см. Техническое Руководство Пищеварение и Кормление [Глава 3]). Глюкоза, необходимая корове в качестве сырья для производства лактозы, синтезируется в печени. В ней основная часть глюкозы образуется в ходе метаболизма пропионовой кислоты, а оставшаяся доля получается в результате распада аминокислот.
Исследования, изучавшие концентрацию глюкозы в крови, попадающей в молочные железы во время лактации, и сравнивавшие ее с концентрацией на выходе из молочных желез, показали, что более 25% глюкозы извлекается из крови во время ее прохождения через ткань молочных желез.
Когда глюкоза попадает в клетки молочных желез, она используется в четырех процессах.
1) Основная часть (около 60-70%) используется для производства лактозы.
Остаток разделяется между следующими процессами:
2) Часть идет на стимуляцию производства белков;
3) Часть преобразуется в глицерин в ходе предварительного этапа производства жира.
4) Часть перерабатывается в ферменты, необходимые для производства жира.
Лактоза является дисахаридом, т.е. веществом, состоящим из двух Сахаров - глюкозы и галактозы.
Соединение одной молекулы галактозы и глюкозы. Соединение одной молекулы глюкозы и галактозы дает молекулу лактозы. Эта связь образуется синтетазой лактозы, которая тем самым является ключевым элементом для синтеза лактозы.
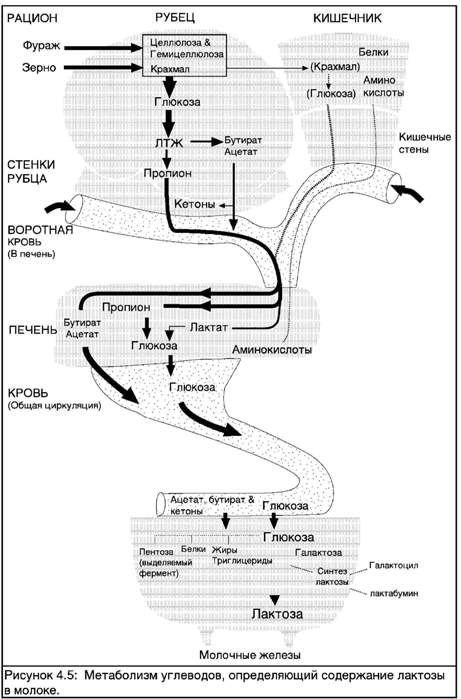
Синтез лактозы в альвеолях вызывает втягивание в них воды. Каждый микрограмм лактозы связывает приблизительно в десять раз больший объем воды. Лактоза является одним из основных веществ, определяющих объем молока, поскольку она определяет половину осмотического давления молока, и тем самым контролирует объем воды. Таким образом синтетаза лактозы чрезвычайно важна для определения количества производимого молока или объема лактации. Синтетаза лактозы образована двумя предшествующими ей субъединицами - ос-лактальбумином и галактозил трансферазой.
Генетические вариации в объеме синтеза сс-лактальбумина могут являться важным индикатором потенциала молочной продуктивности.
СИНТЕЗ МОЛОЧНОГО БЕЛКА
За исключением альбумина и иммуноглобулинов, которые синтезируются вне клеток молочных желез и переносятся к эпителиальным клеткам молочных желез в "готовом" состоянии, белки молока синтезируются секреторными клетками молочных желез из аминокислот, поступающих к ним через кровь. Некоторые из аминокислот называются "незаменимыми аминокислотами". Они должны поступать по кровотоку.
Таблица 4.3: Важные и второстепенные аминокислоты в синтезе молочного протеина*
|
Важные |
Второстепенные |
|
аминокислоты |
аминокислоты |
|
Метионин |
Глютаминовая к-та |
|
Фенилаланин |
Тирозин |
|
Лейцин |
Аспарагин |
|
Треонин |
Орнитин |
|
Лизин |
Аспарагиновая к-та |
|
Аргинин |
Алании |
|
Изолейцин |
Глютамин |
|
Гистидин |
Глицин |
|
Валин |
Цитрулин |
|
|
Серии |
*На основании экспериментов, проведенных на козах.
Остальные "несущественные" аминокислоты могут либо поступать через кровь, либо синтезироваться клетками молочных желез по мере необходимости.
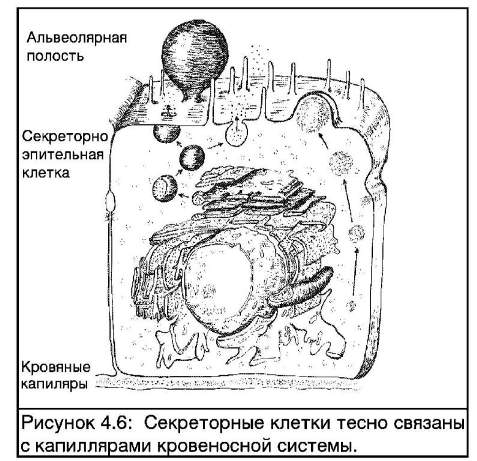
Синтез белка в клетках молочных желез протекает в слоях грубой эндоплазматической сетки (на Рис. 4.6 показано схематичное строение секреторной клетки). Структура белка определятся генетической информацией (ДНК), содержащейся в клетке. По образцу ДНК формируются матрицы РНК, с которых уже в свою очередь копируются последовательности аминокислот, образующих белок. Вакуоли (маленькие пузырьки) в цитоплазме клетки переносят белок к поверхности клетки и выбрасывают его наружу клетки при слиянии вакуоли с клеточной мембраной.
Некоторые белки, присутствующие в молоке, не образованы в клетках молочных желез, а попадают туда из крови. Так альбумин, содержащийся в молоке, синтезируется в печени и его содержание в молоке отражает его концентрацию в сыворотке. Иммуноглобулины попадают в молоко из селезенки и лимфатических узлов через
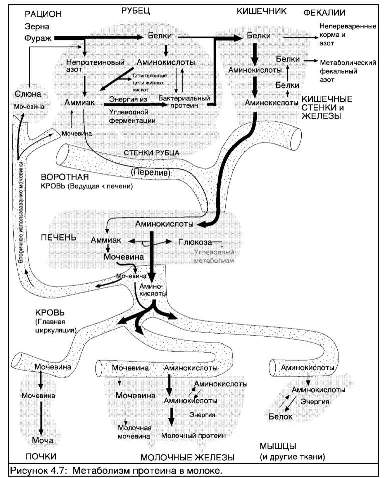
кровь и лимфу. Производящие антитела лимфоциты крови могут присутствовать в молочных железах в раннем периоде лактации. Они могут вносить вклад в производство иммуноглобулина G (IgG), присутствующего в молозиве, но они не наблюдаются в поздней лактации.
СИНТЕЗ МОЛОЧНЫХ ЖИРОВ
Корова получает с кормами только половину объема жиров, присутствующих в молоке. Растительные жирные кислоты, потребляемые в составе рациона, являются сильно ненасыщенными (с недостатком атомов водорода). В желудке происходит добавление атомов водорода (насыщение) до того, как жирные кислоты попадают в кровоток циркулируют там в виде богатых триглицеридом липопротеидов (см."Липидный метаболизм молочных коров"). Эти липопротеиды, богатые триглицеридом, переносятся к клеткам молочных желез; однако они слишком велики, чтобы прямо пройти сквозь клеточную мембрану, и поэтому должны быть разделены на два компонента -глицерин и жирную кислоту - с помощью фермента (липазы липопротеида), присутствующего в стенках капилляров молочных желез. Произведенная в результате жирная кислота может проникать в клетки молочных желез. Исследования, сравнивающие количество богатых триглицеридом липопротеидов в крови на входе в молочные железы и на выходе из них, зарегистрировали резкое падение концентрации триглицерида в результате поглощения жирных кислот в ткани молочных желез, которые затем используются в качестве строительных блоков при синтезе молочного жира.
Некоторые другие "свободные жирные кислоты" могут попадать в молоко напрямую из крови; в них входят жирные кислоты с длинными цепями. Тем не менее, основную часть жирных кислот в молоке составляют кислоты с короткими цепями, содержащими менее 16 атомов углерода и произведенными непосредственно клетками молочных желез.
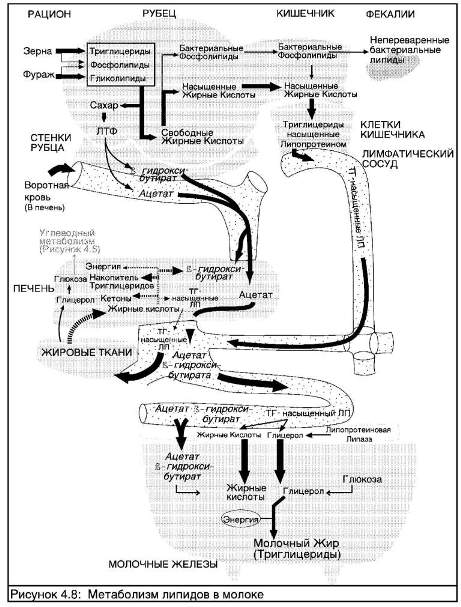
Среди летучих жирных кислот, вырабатывающихся в желудке и попадающих в кровь, присутствуют ацетатная и масляная кислоты Эти две кислоты используются в качестве строительных блоков для синтеза коротких жирных кислот, преобладающих в молоке. Около 17-45% молочного жира строится из уксусной кислоты и около 8- 25% из масляной. Таким образом, ферментация в желудке самым тесным образом связана с производством молока, и Молочный Жир (Триглицериды)
Приблизительно половина жира, содержащегося в молоке, поступает из рациона коровы, остальная часть синтезируется в молочных железах.
эффективная желудочная ферментация является необходимым условием для обеспечения непрерывной поставки летучих жирных кислот. Некоторые побочные продукты уксусной и масляной кислот (так называемые кетоновые тела) также могут быть использованы для построения молочных жиров. Примечательно, что третья летучая жирная кислота, производящаяся в результате желудочной ферментации, пропионовая - используется для синтеза лактозы, но не участвует в процессе образования жира.
Поскольку синтез молочного жира настолько динамичен, изменения в рационе могут менять долю различных жирных кислот в общем составе молочного жира, хотя такие изменения и не влияют на жир, запасаемый в качестве резервов организма.
ПРОИЗВОДСТВО СОЛЕЙ И МИНЕРАЛОВ, СОДЕРЖАЩИХСЯ В МОЛОКЕ
Хотя молоко и содержит множество различных микроминералов, преобладающую роль играет транспортировка через эпителий молочных желез тех минералов, которые важны для правильного развития скелета молодого организма млекопитающего. Среди минералов, задействованных в росте костей, присутствуют кальций, фосфор и, в меньшей степени, магний. Концентрация этих минеральных элементов в молоке существенно (на порядок) больше их содержания в крови. Такое возможно благодаря тому, что ионы кальция и магния в молоке связаны с аминокислотами в молекулах казеина.
Содержание калия, натрия и хлорида в молоке является вторым по значению фактором, определяющим объем воды, попадающей в молоко (путем воздействия на осмотическое давление). Одни дополняют воздействие лактозы, определяющее объем производимого молока.
НАКОПЛЕНИЕ И ВЫСВОБОЖДЕНИЕ МОЛОКА
Весь процесс секреции молока протекает в альвеолярном эпителии. Альвеолярные протоки являются непроницаемыми с кровотоком в отношении обмена веществ. После того, как молоко попадает в молочные протоки, оно стекает в полость, где и накапливается до тех пор, пока корову не начнет сосать теленок или ее не будут доить. По мере того, как молоко "задерживается" и скапливается в вымени, входящие в его состав белки и углеводы накапливаются в альвеолярных клетках, что замедляет темп синтеза. Поэтому частое удаление молока из вымени помогает компонетам молока вытекать из секреторных клеток, тем самым увеличивая выход молока и обеспечивая непрерывное функционирование механизма секреторных клеток.
ЭНЕРГИЯ, НЕОБХОДИМАЯ ДЛЯ ПРОИЗВОДСТВА МОЛОКА
В дополнение в обеспечению сырья для производства компонентов молока, рацион коровы должен обеспечивать достаточное количество энергии для поддержания процессов синтеза, протекающих при секреции молока.
Таблица 4.4 демонстрирует, что наибольшая часть энергии необходима для синтеза жира, тогда как производство лактозы является относительно энергетически экономичным. Если учитывать питательную ценность производимого молока, то можно отметить, что молочные железы очень эффективно расходуют энергию. На каждые 100 Мкал потребляемой коровой энергии, идущей на производство молока, только около 10% используется собственно на процесс производства молока, и около 90% сохраняется в виде энергии, содержащейся в молоке и доступной теленку или другим потребителям молока. Эта величина может быть удвоена, если учесть питательную ценность жира, синтезированного не в молочной железе.
Таблица 4.4: Содержание жирных кислот в триглицеридах молочного жира.
|
Жирные |
Количество атомов |
% |
|
кислоты |
углерода |
|
|
Масляная |
4 |
3,7 |
|
Caproic |
6 |
2,0 |
|
Caprylic |
8 |
1,3 |
|
Capric |
10 |
2,7 |
|
Laurie |
12 |
4,0 |
|
Myristic |
14 |
7,9 |
|
Пальмити- |
16 |
10,7 |
|
новая |
|
|
|
Ситеарино- |
18 |
10,7 |
|
вая |
|
|
|
Арахиновая |
20 |
0,5 |
|
|
—Ненасыщенные— |
|
|
Пальмито- |
16,1 |
1,8 |
|
леиновая |
|
|
|
Олеиновая |
18,1 |
38,3 |
|
Линолевая |
18,2 |
4,7 |
Все жирные кислоты в составе молока содержат четное количество атомов углерода.
Небольшой поправкой, не учтенной в вышеприведенных подсчетах, является энергетическая стоимость переработки пропионовой кислоты в глюкозу (процесс глюконеогенеза) для синтеза лактозы. Этот процесс состоит из нескольких этапов, но можно подсчитать, что для синтеза каждой молекулы глюкозы необходимо около 42 калорий энергии. Эта поправка объясняет разницу между
Таблица 4.5: Энергия, расходуемая в молочной железе на производство 50 кг молока.
|
Компонент |
% |
молей |
ккал/ |
ккал |
общий |
|
|
|
50 |
моль |
общий |
% |
|
|
|
литров |
|
|
|
|
Белки |
4 |
20 |
28 |
560 |
23 |
|
Жиры 1 |
2 |
4 |
455 |
1820 |
73 |
|
(4%хО,5) |
|
|
|
|
|
|
Лактоза |
5 |
7 |
14 |
100 |
4 |
|
Общее |
|
|
|
2480 |
|
|
количество |
|
|
|
ккал |
|
1 Приблизительно половина жира производится в клетках молочных желез, а вторая половина поступает в виде готовых жирных кислот. Затраты энергии на производство половины от полного количества молочного жира включены в приведенный выше расчет.
энергетическими потребностями для производства молока, рассчитанными при определении питательных требований (см. Техническое Руководство Пищеварение и Кормление) и цифрами, приведенными здесь.
Таблица 4.6: Калорийная ценность 50 кг молока
|
Компонент |
Грамм |
ккал/г |
Питательная |
|
|
|
|
ценность |
|
|
|
|
ккал |
|
Белки |
2000 |
4 |
8,000 |
|
Жиры |
1000 |
9 |
9,000 |
|
Лактоза |
2500 |
4 |
10,000 |
|
|
|
|
27,000 |
МЕТАБОЛИЧЕСКИЕ ПРОБЛЕМЫ,
СВЯЗАННЫЕ С ПРОИЗВОДСТВОМ МОЛОКА
Выделение молока наступает внезапно при отеле и возрастает ежедневно. Поэтому для удовлетворения потребностей организма при лактации ежедневно требуется возрастающее количество компонентов, необходимых для производства молока. У животного существует растущая потребность в аминокислотах, жире, глюкозе, кальции и фосфоре. Организм обладает запасами первых двух компонентов, которые он может расходовать для удовлетворения внезапно возросших потребностей, но мобилизация глюкозы и кальция более трудна для организма. Нехватка глюкозы может вызвать кетоз, а недостаток кальция - родильный парез (молочную лихорадку).
Неспособность мобилизовать питательные вещества при началелактации может привести к возникновению метаболических проблем.
КЕТОЗ
Кетоз вызывается главным образом истощением источников глюкозы,
Таблица 4.7: Сравнение затрат энергии на производство молока с его калорийной ценностью.
|
Компоненты |
Энергия синтеза |
Питательная |
Эффектив- |
|
|
|
ценность |
ность |
|
|
ккал (затраты) |
ккал (выход) |
затраты/выход |
|
Белки |
560 |
8000 |
14,3 |
|
Жиры |
1820 |
9000 |
4,9 |
|
Лактоза |
100 |
10,000 |
100,0 |
|
Общее |
2,480 |
27,000 |
10,88 |
|
количество |
|
|
|
циркулирующей в крови. На каждый дополнительный литр молока, производимого коровой, ей необходимо 50 г глюкозы для синтеза лактозы. Поэтому при начале лактации корове нужно быстро увеличить объем потребления углеводов, поддающихся ферментации, и скорость синтеза глюкозы. Если рацион не обеспечивает достаточного количества энергии, готовой к ферментации (см. Техническое Руководство Пищеварение и Кормление), то корова не может произвести достаточное количество пропионовой кислоты, необходимой для синтеза глюкозы. Уровень содержания глюкозы в крови может упасть до половины от нормального уровня. Организм коровы ощущает это как нехватку энергии. Поскольку у нее нет резервов глюкозы, ее организм пытается компенсировать нехватку, мобилизуя резервы жира и протеина. Жир расщепляется с образованием жирных кислот (процесс окисления). Жирные кислоты, как например пропионовая, используются для синтеза глюкозы метаболическим путем, который является обратным нормальному процессу их метаболизма, называемому глюконеогенезисом. Побочным продуктом обратного глюконеогенеза является накопление группы веществ, называемых "кетоновыми телами", по которым и названо заболевание. Присутствие излишка кетоновых тел можно определить по наличию характерного запаха у дыхания коровы. Некоторые аминокислоты, содержащие глюкозу (глюкогенные аминокислоты), также подвергаются расщеплению.
Кетоз обычно начинается между 10-м днем и шестой неделей лактации, но наиболее часто приблизительно через три недели после отела, когда кривая лактации достигает пика. Все коровы обычно испытывают кетоз в некоторой степени, но только у небольшого процента (4-12%) заметны физические проявления. Корова теряет аппетит, ее поведение становится пассивным и наблюдается запор. Риск кетоза возрастает, если коровы была перекомлена (ожирение) перед отелом. Это уменьшает ее аппетит и потребление обычных источников глюкозы, а также устанавливает с самого начала лактации такую структуру обмена жиров, которая приводит к накоплению кетоновых тел.
Наиболее высок риск возникновения кетоза для коров, которые были перекормлены до отела.
Внутривенное введение глюкозы может дать только временное подавление кетоза благодаря повышению содержания глюкозы в крови. Эффективность такого решения проблемы ограничена, т.к. излишнее количество глюкозы удаляется из организма почками. Другие методы, например введение пропиленгликоля, направлены на увеличение содержания пропионовой кислоты в желудке, стимулируя тем самым образование глюкозы.
Кетоз редко заканчивается смертельным исходом; у тех немногочисленных коров, которые умирают, обнаруживается скопление жира в печени. Обычно это состояние самоограничивается, поскольку пониженное потребление кормов
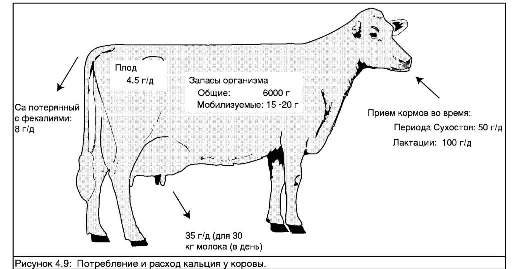
Рисунок 4.9: Потребление и расход кальция у коровы.
ограничивает производство молока и тем самым снижает потребность в глюкозе.
МОЛОЧНАЯ ЛИХОРАДКА ИЛИ РОДИЛЬНЫЙ ПАРЕЗ
Состояние молочной лихорадки наблюдается у только что отелившихся коров. Чуть более одного грамма кальция нужно корове для производства каждого литра молока. Поэтому высокоудойные коровы испытывают внезапную потребность в кальции при начале лактации. Основным источником кальция в организме являются кости, но кальций не может быть мобилизован из них быстро. Медленная мобилизация резервов в костях начинается в качестве реакции на возросшую потребность в кальции. Однако только по достижении десятого дня лактации выделения из костей достигают уровня, дающего существенный вклад в уровень содержания кальция в крови. До наступления этого момента организм коровы зависит от эффективного поступления кальция через желудочно-кишечный тракт. Таким образом, корова подвергается наибольшему риску возникновения родильного пареза в первые десять дней лактации.
Корова подвергается наибольшему риску возникновения молочной лихорадки в первые десять дней лактации.
Дефицит кальция можно заметить по нарушению мышечных функций, т.к. кальций необходим для передачи нервно-мышечных сигналов. Коровы, страдающие молочной лихорадкой, обычно не могут стоять и выказывают признаки слабости мышц, например шейных. Корова дремлет или выглядит сонной. В противоположность названию "молочная лихорадка" температура тела коровы обычно понижена. Родильный парез может оказаться смертельным, если лечение не начато быстро. Внутривенное вливание солей кальция используется для восстановления уровня содержания кальция в крови.
Профилактические меры обеспечивают адаптацию коровы к присутствию кальция в кишечнике к моменту отела.
Это достигается иногда снижением потребления кальция до отела и , скармливанием витамина Д для стимуляции способности усвоения кишечника и мобилизации кальция костей
| ОСНОВНЫЕ ПУНКТЫ Лактация начинается сразу после отела в результате реакции на сложную последовательность гормональных сигналов, которые также инициируют'.. отел. Процессы ферментации, котролирующие синтез лактозы, включаются при отсутствии прогестерона. Производство молока быстро возрастает в течение первых трех-шести недель лактации, а затем медленно спадает. Производство молока прекращается, когда корову больше не доят. Корова должна быть переведена в сухостойное состояние приблизительно за два месяца до следующего отела, чтобы дать молочным железам возможность подготовиться к лактации. Молоко выпускается в протоки молочных желез в результате сокращения миоэпителиальных клеток, окружающих альвеоли. Действие этих клеток контролируется гормоном окситоцином. Все, что корова видит или слышит и что ее беспокоит, может блокировать отдачу молока. Объем лактации может быть подвержен влиянию нескольких факторов, включающих породу, число предыдущих лактации, длину сухостойного периода, месяц отела и несколько факторов содержания, как например доступ к кормам и воде или использование гормона роста. Основным углеводом в молоке является лактоза - дисахарид, синтезированный из глюкозы. Некоторые белки, такие как иммуноглобулины и альбумин, транспортируются к молочным железам в готовом виде. Другие белки, специфичные для процесса лактации, производятся секреторными клетками. Молочные железы также производят около половины количества жира, которое должно содержаться в молоке. Начало интенсивной синтезирующей активности может приводить к метаболичестким дисбалансам при наступлении лактации. Такие нарушения включают в себя энергетические дисбалансы, приводящие к кетозу, и неадекватное снабжение кальцием, вызывающее молочную лихорадку.
|







