
ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ МЛЕКОПИТАЮЩИХ
Для млекопитающих характерен внутренний способ оплодотворения, при котором сперматозоиды при эякуляции достигают яйцеклетки с помощью механизмов положительного хемотаксиса и отрицательного реотаксиса, стереотаксиса. Активно перемещаясь по женским половым путям, половые клетки самца достигают яйцеклетки в маточной трубе, где непосредственно и осуществляется оплодотворение. Для него, в отличие от других классов животных, более характерна моноспермия.
Дробление у млекопитающих начинается с конца первых суток после оплодотворения, продолжается в течение 3-4 суток во время движения зародыша по маточной трубе. Первоначально питание зародыша происходит лишь за счет запасов желтка, накопленных в ово-ците, но уже на поздних стадиях дробления зародыш начинает использовать питательные вещества, получаемые из содержимого секрета, выделенного железистыми клетками яйцевода и матки.
Яйцеклетка у живородящих млекопитающих вторично олиголе-цитальная, почти изолецитальная, в связи с чем дробление у них полное, почти равномерное, асинхронное. Первое и второе дробление проходит меридионально, а в последующем — поперечно. В ходе дробления формируется морула, из нее образуется бластоциста. Обнаружено, что с самого начала имеются клетки с более высокой митотической активностью. Клетки ранней бластулы слабо связаны между собой и могут легко оторваться. Такие автономно лежащие бластомеры полнее могут сформировать новый зародыш. Два зародыша, которые сформировались из бластомеров одной зиготы, имеют идентичный геном и называются однояйцевыми близнецами.
До третьего дробления бластомеры располагаются рыхло, но с вось-мого бластомера адгезивные свойства их мембран увеличиваются, и клетки начинают плотно прилежать друг к другу. Объем межклеточного вещества между клетками резко уменьшается. Определенную роль в этом процессе играет накопление в цитолемме белка увоморулина, способствующего слипанию соседних мембран. Появляются щелевид-ные контакты между бластомерами, обеспечивающие ионообмен между клетками и, следовательно, более активное функциональное взаимодействие.
Со стадии 16-32 бластомера зародыш располагается в нижней трети маточной трубы. В нем можно выделить внутренние и наружные клетки. Внутренние клетки более плотные и обозначаются как темные бластомеры. Клетки с большей митотической активностью имеют высокую вероятность оказаться внутри, но процесс этот не носит полностью детерминированного характера и в какой-то мере случаен. Внутренние бластомеры являются аполярными клетками, тогда как наружные — полярны. Аполярные клетки обозначаются как светлые бластомеры и характеризуются высокой плотностью щелевых контактов и очень активно взаимодействуют между собой. Адгезивные свойства аполяр-ных клеток снижены, а цитоплазма содержит значительное количество органелл. С момента погружения внутрь бластулы их митотическая активность падает. Полярные же клетки отличаются тем, что их наружная, покрывающая бластулу поверхность неровная и содержит значительное количество микроворсинок. Межклеточные поверхности имеют выраженную склонность к прилипанию, на них нет микроворсинок, а цитоплазма бедна органеллами.
Зрелая (свободная) бластоциста млекопитающего, достигнув полости матки, освобождается от блестящей оболочки и питается за счет выделяемого маточных желез. Реснитчатый эпителий эндометрия матки перемещает ее по ходу матки. Зародыш состоит из трофобласта (зак-чатка зародыша) и эмбриобласта (закладки эпителия хориона). Видна полость, заполненная жидкостью, — бластоцель. После разрушения прозрачной оболочки зародыш может непосредственно контактировать с эпителием эндометрия матки. Взаимодействие рецепторов трофобласта зародыша с покровными эпителиоцитами эндометрия сопровождается прилипанием зародыша к стенке матки. Это явление назы-вается адгезией. Прилипший зародыш раздвигает эпителиоциты и начинает погружаться в стенку матки. Погружение зародыша в эндометрий обозначается термином имплантация. Трофобласт начинает бурно развиваться, выделяет протеолитические ферменты, частично разрушающие стенку матки. Клетки трофобласта усиленно делятся, и поверхность зародыша становится неровной, покрываясь множеством ворсинок. Эпителий трофобласта дает начало формирующемуся эпителию ворсинок хориона. Эндометрий с момента погружения зародыша в стенку матки бурно разрастается, активно развиваются железы матки и активизируется выделение ими секрета, в стенке эндометрия развиваются сосуды и децидуальные клетки. Функциональный слой эндометрия формирует основу отпадающей (децидуальной) оболочки стенки матки беременного животного.
Имплантация обычно происходит одновременно с началом первой стадии гаструляции. Имплантация — процесс внедрения (у хищных, приматов, грызунов) или прикрепления (копытные, свиньи) зародыша в стенке матки. Трофобласт зародыша перед адгезией (прилипанием) растворяет прозрачную зону за счет образования им гидролитических ферментов и прилипает к эпителию. В последующем у некото-рых млекопитающих (приматы, хищные) происходит дальнейшее внедрение в эндометрий с погружением зародыша в его толщу. У крупного рогатого скота прикрепление происходит на 17-е сутки.
Начиная с первой стадии гаструляции происходят и другие события. От внутренней зародышевой массы (эмбриобласта) в бластоцель мигрируют клетки (см. рис. 1), которые в последующем служат материалом для внезародышевой энтодермы. Эти клетки образуют стенку желточного пузырька. При этом клетки внезародышевой энтодермы
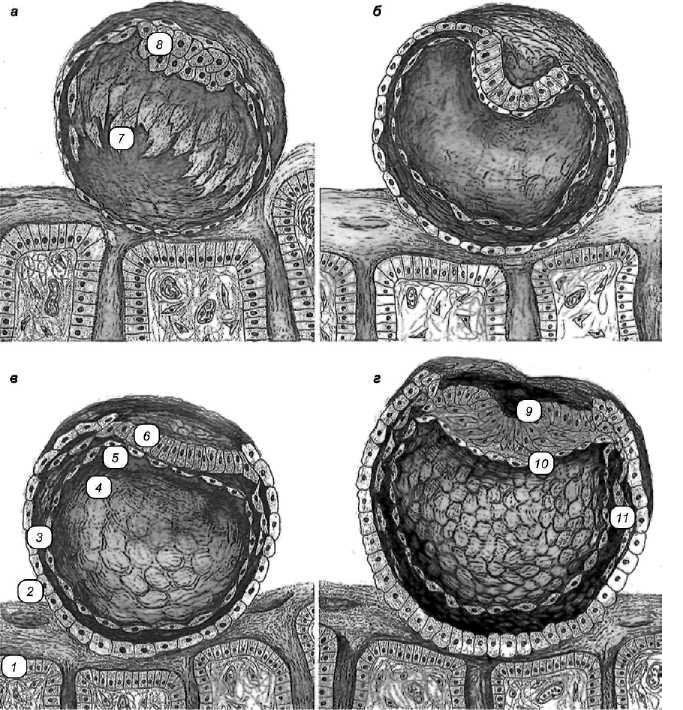
Рис. 1 Гаструляция у млекопитающих (сельскохозяйственных животных):
а — бластоциста; б — формирование двух зародышевых листков; в — образование внезаро-дышевой мезодермы; г — образование зародышевой мезодермы. 1 — эндометрий матки; 2 — трофобласт; 3 — внезародышевая энтодерма желточного мешка; 4 — полость зародышевого мешка; 5— гипобласт; 6— энибласт; 7 — бластоцель; 8— эмбриобласт; 9— первичная полоска; 10, 11 — зародышевая и внезародышевая мезодерма.
тесно прилежат к трофобласту. Трофобласт над эмбриобластом у сельскохозяйственных животных истончается, и последний начинает взаимодействовать с полостью матки. Эмбриобласт путем деламинации начинает подразделяться на два зародышевых листка — поверхностно лежащий эпибласт и прилежащий к бластоцели гипобласт. Гипобласт составлен уплощенными и рыхло расположенными клетками, тогда как эпибласт отличается тесно связанными клеточными структурами. К этому сроку виден желточный пузырек. Зародыш раздвигает трофобласт и уплощается, приобретая форму диска, приближаясь по форме к зародышевому диску птиц.
Вторая стадия гаструляции, по механизму напоминает подобный процесс у птиц, и в ее основе лежит иммиграция. В результате перемещения клеток в эпибласте закладывается первичная полоска и первичный узелок. От первичной полоски в латеральном направлении мигрируют клетки — предшественники зародышевой мезодермы. В результате образуются три зародышевых листка — эктодерма, мезодерма и энтодерма. Видна также внезародышевая мезодерма и внезаро-дышевая энтодерма. В результате перемещения материала, примыкаю-щего к первичному узелку (как и у птиц), формируется хорда. Она индуцирует образование нервной пластинки, а затем нервного желобка и нервного гребня. По бокам от них видна зародышевая эктодерма. Вентральнее лежит зародышевая и внезародышевая энтодерма. В зародышевой мезодерме появляются скопления клеток: тело сомита и его ножка. Латеральнее ножек сомита располагается целом. Туловищная складка у млекопитающих появляется довольно рано и отделяет формирующийся зародыш от внезародышевых органов. Удлинение зародыша и появление туловищной складки обеспечивают закладку первичной кишки.
Внезародышевая эктодерма, трофобласт и внезародышевая мезодерма составляют основу амниотической складки по бокам от формирующегося зародыша в ранние сроки гаструляции сельскохозяйственных животных. В ходе гаструляции свободные края амниотической складки увеличиваются и направляются друг к другу, сливаются между собой, образуя амниотическую полость. По мере развития туловищной складки зародыш оказывается в полости амниона, заполненной амниотической жидкостью. Стенка амниона и лежащего над ним хориона участвует в формировании плодных оболочек. Плодные оболочки защищают зародыш от инфекции, механических повреждений, воздействия других внешних факторов, обеспечивают существование зародыша в жидкой среде вплоть до рождения. В зону между внезаро-дышевой эктодермой и трофобластом рыхло прорастают клетки внутренней зародышевой массы, образуя внезародышевую мезодерму.
Эпителий амниона имеет различное строение в зависимости от вида животного, сроков его развития. Его строение может быть от однослой-ного призматического до однослойного плоского эпителия. Над плацентой эпителиоциты призматической формы. Соединительнотканная оболочка амниона образована несколькими слоями плотной волокнистой соединительной ткани. Амнион связан с хорионом прослойкой рыхлой соединительной ткани, содержащей кровеносные сосуды.
Формирование желточного пузырька у млекопитающих происходит в ходе гаструляции. Процесс начинается с миграции части клеток внутренней зародышевой массы в полость бластоцеля. Рыхло лежащие клетки частично заполняют бластоцель и образуют пузырек вен-тральнее гипобласта. От гипобласта мигрируют клетки, выстилая по-лость желточного пузырька. В отличие от животных с полилециталь-ными яйцеклетками, желточный пузырек млекопитающих не содержит трофических включений. В мезенхиме стенки желточного мешка дифференцируются в островки кроветворения и кровеносные сосуды. Кроме этого, в его стенке развиваются гонобласты (родоначальники половых клеток). Желточный мешок у млекопитающих быстро подвергается инволюции и присоединяется к пупочному канатику.
Аллантоис, как и у птиц, образуется отделением от желточного пузырька в хвостовом конце зародыша выпячивания. Аллантоис у млекопитающих, в отличие от аллантоиса птиц, небольших размеров. Его стенка составлена внезародышевой энтодермой и мезенхимой. В ходе развития мезенхима дифференцируется в слизистую соединительную ткань. Стенка аллантоиса служит основой пупочного канатика, в стенку которого прорастают кровеносные сосуды (закладки пупочных артерий и вен).
Пупочный канатик во второй половине внутриутробного развития представляет собой эластичный тяж, связывающий плод с плацентой. Основу пупочного кантика образует слизистая соединительная ткань. В ней располагаются пупочные артерии и вены. В его толще можно найти остатки полости аллантоиса и желточного пузыря. Поверхность пупочного канатика покрыта однослойным плоским эпителием амниона.
Закладка хориона происходит в процессе имплантации. Его эпителий образуется из клеток трофобласта. Бурно развиваясь, клетки трофобласта из одного слоя могут сформировать несколько слоев. Поверхностные слои не контактируют с производными эмбриобласта и теряют способность к митотическому делению. Эти клетки могут сформировать симпласт. В результате в эпителии хориона можно выделить два слоя: внутренний клеточный цитотрофобласт и наружный некле-точный симпластотрофобласт, или синцитиотрофобласт. Эпителий тро-фобласта образован клетками призматической формы с обилием рибосом и полисом, других органелл. Симпластотрофобласт состоит из многоядерных образований (симплатов) с базофильно окрашенной
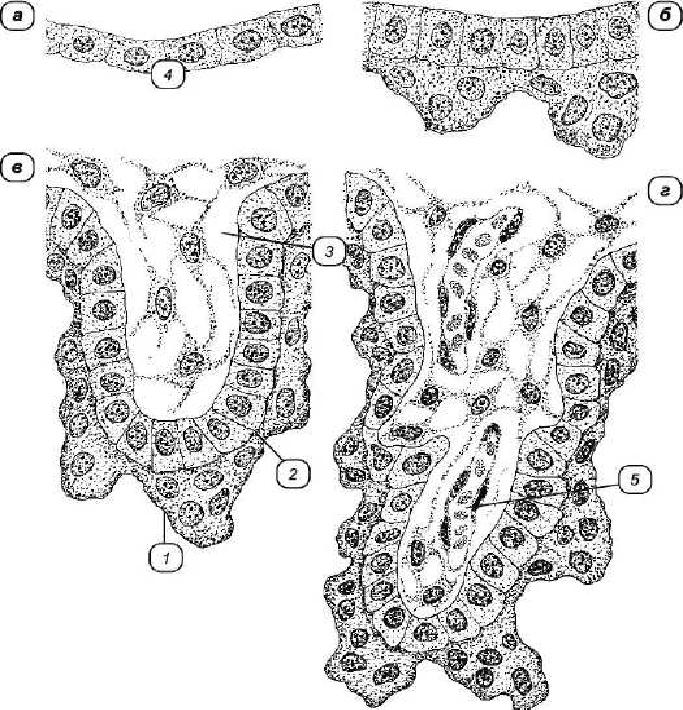
Рис. 2 Формирование ворсин хориона:
а — трофобластический эпителий; б — первичная ворсина хориона; в — вторичная ворсина; г— третичная ворсина. 1— симпластотрофобласт; 2 — цитотрофобласт; 3 — мепенхима; 4 — ппителий трофобласта; 5 — кровеносный сосуд.
цитоплазмой, с обилием микроворсинок на апикальной поверхности синтетических органелл, митохондрий. Симпластотрофобласт не выводит на своей апикальной поверхности антигенных комплексов, предотвращая реакции отторжения зародыша организмом матери.
Хорион, образует ворсинки, покрывающие всю поверхность плодного пузыря, которые вначале состоят только из эпителия хориона и называются первичными ворсинками. В последующем в толщу ворсинок проникают прослойки внезародышевой соединительной ткани. Такие ворсинки называются вторичными. Третичные ворсинки содержат кровеносные сосуды. Процесс формирования кровообращения в хорионе называется процессом плацентации. Процесс плацентации является критическим периодом в развитии млекопитающих (рис. 2).
Хорион, связанный со стенкой децидуальной оболочки, отличается сильным развитием ворсинок и называется ворсинчатым. Участок со слабым развитием ворсинок обозначается как гладкий хорион. Ворсинчатый хорион участвует в образовании плаценты.
Плацента — сложная структура, состоящая из зародышевых и материнских элементов. Со стороны зародыша в нее входит хорион, а со стороны матера — децидуальная оболочка матки (отпадающая оболочка, которая развивается из функционального слоя эндометрия).
Для плаценты характерны следующие функции.
1. Барьерная функция. Плацента обеспечивает избирательную проницаемость веществ. Через нее не проникают многие высокомолекулярные вещества, но легко поступают мономеры, вода, ионы, жирорастворимые вещества. Ведущим элементом барьера является эпителий хориона.
2. Трофическая функция. За счет плаценты в организм зародыша поступают все необходимые для его развития вещества.
3. Газообменная функция. Через плаценту осуществляется газообмен, поступает кислород, выделяется углекислый газ. Для зародыша млекопитающего плацента является своеобразным органом дыхания.
4. Экскреторная функция. Через плаценту организм зародыша выделяет продукты своего метаболизма. Таким образом плацента осуществляет детоксикационную функцию, поддерживает кислотно-щелочное равновесие.
5. Эндокринная функция. Плацента выделяет большое число гормонов, регулирующих течение беременности, метаболические процессы матери и плода. Известна большая роль хорионического гонадотропина, образуемого эпителием ворсин хориона. Этот гормон стимулирует гормональную активность лютеоцитов желтого тела, подавляет лимфоциты матери, предотвращая реакции отторжения плода. Кроме этого, хорионический гонадотропин оказывает значительное адаптогенное действие. Плацента также способна к образованию прогестерона и релаксина, эстрогенов. Соматоммамотропин плаценты стимулирует рост секреторных отделов молочной железы и иногда называется плацентарным лактогеном. Значима роль трансферрина плаценты, который участвует в эмбриональном гемопоэзе, связывая железо. Плацента также способна к образованию кортиколиберина; тропных гормонов, аналогичных гормонам гипофиза (тиротропину, адренокортикотропину, меланотроину); андрогенов, некоторых факторов роста.
6. Иммунологическая функция. Через плаценту в зародыш проникают иммуноглобулины матери, защищая плод от проникновения в него инфекции.
7. Депонирующая функция. В плаценте накапливаются многие необходимые для организма зародыша вещества — минеральные соли, жирорастворимые витамины, витамин С.
8. Гемохориальная плацента регулирует процессы свертывания крови и фибринолиза, в частности предотвращет тромбообразование в сосудистых лакунах плаценты.
Зародышевая часть плаценты представлена хориальной пластиной или пластинами. Ее основа образована рыхлой волокнистой соединительной тканью, в которой располагаются многочисленные кровеносные сосуды. От хориальной пластины в сторону матки направляются многочисленные ворсины. В соединительнотканной основе ворсин лежат мелкие кровеносные сосуды, вплоть до капилляров. Близ сосудов можно найти многочисленные макрофаги (клетки Кащенко-Гофбауера). Хориальную пластину и ворсины снаружи выстилает цитотрофобласт и симпластотрофобласт.
Материнская часть плаценты сформирована децидуальной оболочкой. В области плаценты децидуальная оболочка матки сильно развита. Ее строение различно в зависимости от типа плаценты. Для всех децидуальных оболочек плацент характерно обилие кровеносных сосудов. В соединительной ткани можно найти крупные клетки округлой формы с оксифильно окрашенной цитоплазмой и обилием гликогена. Это децидуальные клетки. Они контролируют развитие трофобласта, местные иммунные реакции, свертывание крови и другие функции.
Десмохориальная, множественная, гистотрофная плацента связывает ворсинки хориона с соединительной тканью децидуальной оболочки. Имеется множество локальных участков, связывающих зародыш с маткой, — множественная плацента (рис. 3). Участки с развитыми ворсинами хориона обозначаются как котиледоны. Соот-ветствующие участки эндометрия матки утолщены и носят название карункулов. В сравнении с эпителиохориальной плацентой она устроена более сложно. Отличия заключаются в том, что ворсинки хориона ветвятся. Наряду с эпителием — цитотрофобластом, имеющим клеточное строение, появляются участки с многоядерными структурами (симпластом) — синцитиотрофобластом (симпластотрофобластом). В участках, где ворсинки взаимодействуют с децидуальной оболочкой, которая частично теряет эпителий, ворсинки хориона прямо взаимодействуют с соединительной тканью.
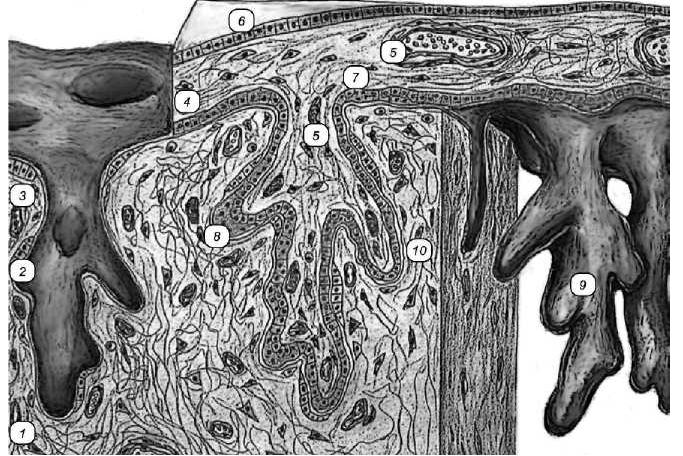
Рве. 3 Десмохориальная плацента:
1 — собственная пластинка децидуальной оболочки матки; 2 — остатки эпителия матки; 3 — кровеносный сосуд децидуальной оболочки; 4 — пластинка хориона; 5 — кровеносные сосуды хориона; 6— эпителий амниона; 7— цитотрофобласт; 8— симпластотрофобласт; 9 — ворсина хориона; 10 — децидуальная клетка.
Подобная плацента характерна для мелких жвачных животных и коров, но это мнение не разделяется некоторыми авторами. Тип питания в такой плаценте близок к типу питания в эпителиохориальной плаценте и также является гистиотрофным.
Внутриутробное развитие большинства сельскохозяйственных животных (крупный и мелкий рогатый скот, лошади, верблюды, свиньи) сопровождается рождением зрелородящихся детенышей.
У крупного рогатого скота продолжительность внутриутробного развития составляет 9 месяцев. Зародышевый (эмбриональный) период охватывает первые 34 суток. Стадия дробления занимает 1-7 суток, стадия питания и дыхания с помощью трофобласта — 8-9 суток. Трофобласт к этим срокам разрастается, приобретает способность к перевариванию выделяемого маточных желез. На этой стадии зародыш начинает увеличиваться в размерах, в нем образуются зароды-шевые листки, осевые зачатки органов, формируется амнион и желточный мешок. К концу этого срока у коровы зародыш прилипает к эндометрию матки.
Стадия питания и дыхания посредством сосудов желточного мешка занимает 20-23 суток. К этому сроку формируются сосуды и кровяные островки желточного мешка, происходит закладка сердца и образование примитивного сосудистого русла. Стадия предполагает формирование закладок многих основных органов (нервной трубки и мозговых пузырей, закладку глаза и внутреннего уха, первичной почки, печени), начинают формироваться сомиты.
Стадия питания и дыхания с помощью сосудов аллантоиса занимает 24-34 суток. В данные сроки происходят процессы плацента-ции и осуществляется устойчивая связь между эмбрионом и хорионом. Этому сопутствует образование основных органов и систем зародыша, происходит миграция нейробластов в закладки нервных узлов, образуются закладки нервов, завершается образование туловищных сомитов, образуются ластоподобные грудные и тазовые конечности.
Предплодный период у коров занимает 35-60 суток. В ранний предплодный период (35-50 суток) бурно развивается хорион, увеличивается число и происходит васкуляризация котиледонов, что значительно улучшает трофическое обеспечение зародыша. Улучшение трофики поддерживается за счет анатомического формирования стенок и закладок клапанного аппарата сердца, а также дуги аорты и закладок магистральных сосудов. Улучшение трофического обеспечения сопровождается формированием хрящевых закла-док скелета.
Поздний предплодный период у крупного рогатого скота предполагает сроки 50-60 суток. Предплод формирует все основные закладки внутренних органов. Заканчивается индифферентная стадия полового развития, и появляются первые половые различия. Появление иннервируемых мышечных волокон позволяет предплоду к концу этой стадии осуществлять первые примитивные движения.
Плодный период занимает 61-270 суток после оплодотворения. В ранний плодный период (61-120 суток) происходит дифференци-ровка клеточных структур, формирующих органы. В нервной ткани появляются юные нейроны (на уровне спинного и ствола головного мозга). Бурно формируются терминальные отделы мозга. Легкое напоминает сложную, разветвленную железу. Образуются внутриор-ганные железы многих полых органов. В кишечнике появляются крипты и ворсинки. Дифференцируются паренхиматозные элементы селезенки, красного костного мозга. Начинает функционировать вторичная почка.
Поздний плодный период (5-9 месяцы) предполагает тонкие молекулярные и клеточные механизмы дифференцировки органов плода. У зрелородящихся животных наблюдается достаточная степень дифференцированности нервного и опорно-двигательного аппарата, что позволяет животному проявлять высокую двигательную активность. Видны породные и индивидуальные признаки зародыша.
Васильев Ю. Г., Трошин Е. И. "Цитология. Гистология. Эмбриология" - Спб: Лань, 2009







