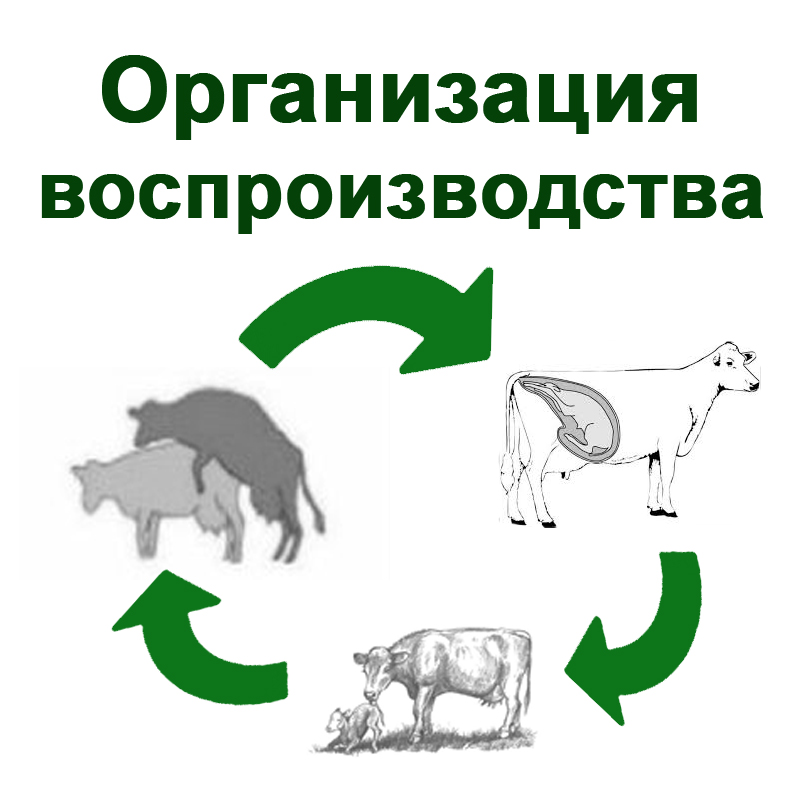

Половой цикл самок
Половой цикл самок - способность неоплодотворенной самки приходить в состояние половой охоты (либидо) через определенные промежутки времени. Он появляет.Г наступаешь половой зреаости у разных видов животных в разное время и повторяется в строго с определенной периодичности до старости У таких животных как коровы, свиньи, Ко0ь,ль половые никлы ритмичны, повторяются со времени наступления половой зрелости до климактерического периода и отсутствуют только в период беременности. Животных у которых-в течение года половой цикл повторяется много раз в определенном ритме, называют полицикличными. Половой цикл полицикличных животных характеризуется сравнительно одинаковыми и короткими фазами.
У некоторых видов животных (собаки, кошки и большинство диких животных) в течение года наблюдается один или два, редко три половых цикла. Таких животных называют моноцикличными или дицикличными. Характерной особенностью полового цикла моноцикличных животных является длительность полового покоя. У других видов животных (овцы, козы, верблюды, буйволы) ритмичность полового цикла отмечается только в течение определенного периода времени, чаще это бывает весной или осенью. Между периодами половой активности наблюдается длительный период полового покоя. Такие животные относятся к полиэстричным, но с половым сезоном (или условно полиэстричные) Половой сезон - время, в течение которого животные находятся в состоянии половой активности, Продолжительность полового цикла у разных животных неодинакова: у коров средняя продолжительность цикла - 18-21 день (с колебаниями - 12-30 дней), у овец - 17 дней (14-19), у свиней - 19-21 день (11-30), у лошадей - 20-22 (10-36).
Половой цикл самок можно разделить на 4 стадии: проэструс (перед охотой), эструс (половая охота), метаэструс (после охоты) и диэструс (не в охоте). По нервным процесса половой цикл делят на 3 стадии: возбуждение, торможение и уравновешивание.
Стадия возбуждения - в кровь из гипофиза по команде гипоталамуса (РГ-ФСГ) выбрасывается ФСГ - начинают созревать фолликулы в яичниках, концентрация фолликулярных гормонов в крови (эстрон, эстриол, эстродиол) увеличивается. Под влиянием эстрогенных гормонов кровеносные сосуды, питающие аппарат размножения, расширяются; усиливается рост клеток в половых органах самок, особенно слизистой оболочки матки и яйцеводов; слизистая оболочка яйцеводов, матки и влагалища становится более красной, отечной; наблюдается попеременное сокращение и расслабление мышечных слоев матки ( рогов, тела и шейки), Вульва также становится отечной, покрасневшей, припухшей. Секреция слизистой оболочки матки, шейки матки, преддверия влагалища значительно усиливается Слизь в канале шейки матки разжижается и вытекает. Все эти изменения функций органов размножения, особенно слизистой оболочки и ее желез, называют течкой - состоянием подготовки животного к оплодотворению и плодоношению.
В это время под влиянием эстрогенов, попадающих с током крови во все ткани, повышается возбудимость нервной системы, усиливается половое возбуждение самки, она становится более подвижной, беспокойной, иногда отказывается от корма. Самка допускает к себе самца (рефлекс неподвижности) и сама стремится к нему, изгибает спину, настораживает уши. Максимальное возбуждение нервных половых центров, характеризующееся готовностью спариваться, называется половой охотой. Это период оптимального времени для естественного и искусственного осеменения. Течка у коров длится в среднем 30 часов, у овец - 37 часов, у свиней - 3-4 дня. Половая охота у самок наступает значительно позже течки. У коров она происходит через 15-20 часов после начала течки и продолжается 17-20 часов, у овец либидо длится 37 часов, у свиней - 48-60 часов.
В конце полового цикла уровень ЛГ, определяющий овуляцию, под влиянием гонадотропина релизинг-гормона быстро увеличивается. В конце первых и начале вторых суток от начала либидо у коров происходит овуляция. У овец, свиней овуляция обычно происходит через 20-40 часов от начала охоты. Овуляция - это разрыв фолликула и выход яйцеклетки из фолликула. У самок сельскохозяйственных животных овуляция происходит спонтанно, т.е.
независимо от встречи самки с самцом. На месте разорвавшегося фолликула формируется и развивается желтое тело - временная половая железа, которая вырабатывает прогестерон и выделяет его в кровяное русло. Реализация ЛГ обусловливается эстродиолом, уровень которого быстро повышается в крови и за 25 часов до охоты, определяя точное время овуляции. При отсутствии беременности количество ЛГ в крови снижается. Уровень прогестерона после 17 дня быстро снижается и одновременно повышается содержание ЛГ и эстродиола. Опыты по пересадке матки и ее денервации подтверждают предположение о передаче сигналов гуморально. Видимо, импульс из матки в яичник на рассасывание желтого тела также осуществляется гуморальным путем. Вещество, вырабатываемое в матке и во влагалище для рассасывания желтого тела, оказывало лютеолитический эффект, почему и названное "лютеолизином матки" представляет собой простагландин-ПГФ
Стадия торможения (метаэструс). После овуляции половая активность самки постепенно угасает. На месте разрыва оболочки яичника и сосудов в опустевшую полость фолликула происходит кровоизлияние. В это время особое значение имеет третий гонадотропный гормон передней доли гипофиза - пролактин или лютеотрофный гормон. Это название указывает на его двоякую функцию. С одной стороны, он подготавливает молочную железу к образованию молока, а с другой - задерживает существование желтых тел в яичнике в том случае, если произошло оплодотворение. В отношении этого гормона еще много неясного, но. по-видимому, справедлива та точка зрения, что для выделения его из гипофиза нужны нервные импульсы, идущие с рецепторов матки. При отсутствии этих стимулов в тех случаях, когда этот гормон не выделяется, желтые тела в яичнике подвергаются дегенерации и через некоторое время наступает новая либидо: Если же головной мозг, по-видимому подбугорье, получает стимулы от плода, находящегося в матке, то подбугорье побуждает гипофиз к выделению лютеотропного гормона, что имеет следствием задержку желтых тел в яичнике и выработку ими прогестерона. Зависимость выделения лютеотропного гормона от рецепции матки доказывается опытами, в которых искусственное введение стеклянных бусинок в матку овцы вызвало задержку дегенерации желтых тел, а перерезка нервов, идущих от матки, делала невозможной имплантацию зародыша. Если оплодотворение не произошло, то рост и секрецию половых органов постепенно прекращается, желтое тело под влиянием лютеолитического фактора матки (ПГ<£>2 ) быстро регрессирует.
Стадия уравновешивания (диэструс). В это время происходит процесс инволюции половой системы, т.е. возвращение ее в состояние до течки и охоты. Через 12-15 дней после овуляции при отсутствии беременности выделение прогестерона постепенно прекращается, клетки желтого тела постепенно дегенерируют, вследствие лютеолитической функции матки активизируются процессы созревания фолликулов. Устранение обратной отрицательной связи влияния прогестерона позволяет гипофизу активно секретировать гонадотропин (ФСГ), под контролем которого созревают один или несколько граафовых фолликулов, выделяя увеличивающиеся количества эстрогенов и стимулируя изменения, которые предшествуют охоте и овуляции.
Половой цикл, таким образом, состоит из чередования относительно длительной фазы желтого тела, в течение которой желтое тело вырабатывает прогестерон, что является основным процессом, происходящим в яичнике и непродолжительной фолликулярной фазы, во время которой один или несколько зрелых фолликулов вырабатывают эстрогены, и этот вид секреции в течение фолликулярной фазы является преобладающим. С точки зрения природы для поддержания популяции данного вида все половозрелые самки должны быть либо беременными, либо в состоянии лактации, а при неудачной попытке достигнуть этого состояния животное приспосабливается так, что процессы, направленные для успешного спаривания, повторяются. Однако, у некоторых видов имеются периоды овариальной бездейственности: у моноцикличных и сезонно полиэстричных животных стадия полового уравновешивания между сезонами называется анефродизией. Описанные четыре стадии состояния самок на основе вагинальной гистологии широко использовались много лет назад до применения прямых способов измерения концентрации гормонов в плазме крови, и они характерны чаще всего мелким лабораторным животным, а не крупным сельскохозяйственным видам. Хотя у сельскохозяйственных животных в течение полового цикла могут быть установлены изменения в структуре вагинальных мазков, они не точны и порой трудноразличимы. Четкое выявление периода самой охоты, что в большей степени интересует животноводов и службу искусственного осеменения, и четкое выявление оптимального периода оплодотворения, особенно с точки зрения поведения, возможны только в присутствии других животных.
Влияние гипофизарных гормонов на половую функцию и качественные изменения в ней под действием каждого из гормонов показаны в таблице 2.
Прежде чем обсуждать примеры гормональной секреции в половом цикле, важно отметить, что хотя овуляция и является «самопроизвольным» явлением во время полового цикла у коров, овец, свиней и лошадей, но это относится не ко всем видам. У таких животных, как кролик, кошка, африканский хорек механизм гонадотропной секреции гипофиза, которая вызывает овуляцию, требует для ее осуществления стимула при спаривании, и таким образм, устанавливается обратная связь между попаданием в половой тракт самки живчиков и выделением овулировавших яйцеклеток. Таким образом, индуцированная овуляция требует афферантных нервных компонентов, исходящих от вагинальной и цервикальной стимуляции до того как будет вызван разрыв фолликула, в то время как у сельскохозяйственных животных овуляция «самопроизвольная», и коитус не нужен для того чтобы спровоцировать предовуляторную волну гонадотропина: его выделение происходит' благодаря влиянию положительной обратной связи, увеличивающей концентрацию эстрогенов из созревших фолликулов. Имеюся данные что при повышенном половом возбуждении в присутствии самца овуляция может ускориться.
|
Место |
|
|
Объект |
Эффект |
|
образования |
|
|
воздей- |
|
|
гормонов |
общее |
специф. |
ствия |
|
|
Щитови- |
|
Тироксин |
Самки |
Способствует поддержанию гипофизармо- |
|
ная железа |
|
гонадального кзаимоотношения и косвенно связан с |
||
|
|
|
воспроизводительной функцией в целом. |
||
|
Передняя |
|
Стимулирует и контролирует развитие фолликулов. |
||
|
доля |
Гонадотро |
Фолликуло |
Яичник |
Контролирует секрецию эстрогенов в яичнике. |
|
гипофиза |
п-ный |
стимулирую- |
||
|
|
гормон |
щий гормон |
||
|
(ФСГ) |
|
Вызывает овуляцию, развитие желтого тела |
||
|
Лютеинизирую |
|
|
||
|
-щий гормон |
Яичники, |
Вызывает секрецию молока и поддерживает
|
||
|
(ЛГ) |
молочная |
лактацию. Стимулирует ЛГ и развитие желтого тела |
||
|
Задняя |
Лютеотро |
Пролактин |
железа |
и образование прогестерона |
|
доля |
п-ный |
|
|
|
|
гипофиза |
гормон |
Матка, |
Стимулирует сокращение матки в период охоты, |
|
|
|
|
молочная |
родов и инволюции гениталий. Вызывает молокоотдачу. |
|
|
|
Окситоцин
|
железа |
|
|
Яичники |
|
Эстродиол, |
Матка и |
Вызывает
течку и охоту На последних стадиях |
|
||||
|
(фолликулярный |
Эстроген |
эстрон и |
молочная |
беременности
обусловливает развитие молочной |
|
||||
|
|
|
четриол |
железа |
железы
и сенсибилизирует машу к окситошну, |
|
||||
|
эпителий и |
|
|
|
регулирует время
овуляции. |
|
||||
|
интерсти- |
|
|
|
|
|
||||
|
циальные |
|
|
|
|
|
||||
|
клетки) Яичники |
|
|
Гипофиз, |
Тормозит
образование ФСГ и угнетает действие |
|
||||
|
Яичники (желтое |
|
Прогестерон |
матка, |
окекгоцина
десенсибилизацией к нему мускульных |
|
||||
|
тело) |
|
|
молочная |
волокон
матки. Вызывает развитие слизистой матки |
|
||||
|
|
|
|
железа |
и секрецию маточных желез, обеспечивает нщвшю |
|
||||
|
|
|
|
|
зиготы. |
|
||||
|
|
|
|
|
При
беременности способствует
дальнейшему |
|
||||
|
|
|
Пролактин |
|
развитию желез вымени, в конце беременности |
|
||||
|
|
|
|
|
подготавливает
молочную железу к
началу |
|
||||
|
|
|
|
|
лактации. |
|
||||
|
|
|
|
|
В конце беременности расслабляет связки таза. |
|
||||
|
|
|
Релаксин |
|
|
|
||||
|
|
|
|
|
В конце беременности стимулирует развитие |
|
||||
|
Плацента |
|
Прогестерон, |
Связки |
молочной
железы |
|
||||
|
|
|
эстроген |
таза |
|
|
||||
|
|
|
|
Гипофиз, |
|
|
||||
|
|
|
|
матка, |
|
|
||||
|
|
|
|
молочная |
|
|
||||
|
|
|
|
железа |
|
|
||||
|
Самцы |
|||||||||
|
Передняя |
Гоиадотр |
Гамето кинети- |
Семеини |
Контролирует развитие извитых канальцев в |
|||||
|
доля |
ОП-НЫЙ |
ческий (ФСГ) |
-ки |
семенниках Стимулирует сперматогенез. |
|||||
|
гипофиза |
гормон |
Гормон, сти- |
|
Контролирует развитие ицтерстиииальных |
|||||
|
|
|
мулирующий |
Семенни |
клеток в семенниках. Стимулирует секрецию |
|||||
|
|
|
интерстициаль |
-ки |
тестостерона в семенниках. |
|||||
|
|
|
-:1ые клетки |
|
|
|||||
|
|
|
(ГСИК или |
|
|
|||||
|
|
|
ЛГ) |
|
|
|||||
|
Семенники |
Андроге |
Тестостерон |
|
Влияет на развитие вторичных половых |
|||||
|
|
н |
|
|
признаков самцов и способствует |
|||||
|
|
|
|
|
проявлению полового влечения (либидо) |
|||||
Эндокринология полового цикла Лютеотропному гормону принадлежит более важна* роль, чем фолликулостимулирующему в регулировании времени овуляции, однако ФСГ имсеп не меньшее значение в развитии граафова фолликула в последних стадиях полового цикла Следует отметить, что предовуляторный пик проявляется незадолго до начала охоты у коров овец и свиней и неизменно противостоит соответствующему ему. но менее выраженному пик ФСГ. Одна из интерпретаций этого явления состоит в том, что существует общи гипоталамический релизикг-гормон для этих двух гонадотропинов. При определении начал полового цикла обычно обозначают первый день охоты как нулевой день (день О) следовательно, овуляция как правило, наступает в 1-й день полового цикла у коров овец исвиней
У коров фаза желтого тела занимает приблизительно 17 дней, продолжительность либидо - 16 часов, этот восприимчивый период, как правило, короче зимой или в самый напряженный период лактации Овуляция происходит через 8-12 часов после окончания охоты или через 25- 30 часов после предовуляторнон волны гонадотропина. когда диаметр полност сформированного фолликула достигает 16-18 мм Концентрация прогестерона периферической крови при приближении охоты составляет менее 1,0 нг/мл"' и до 5-го дн заметно не повышается, затем устойчиво возрастает к 16-17-му дню до 5,4 нг/мл"' во врем фазы желтого тела цикла и достигает пикового значения около 6-7 нг/мл в конце фазы желтого тела. В то время как желтое тело может иметь 2 см в диаметре и более, у циклирующего животного наблюдается снижение концентрации прогестерона Начало снижения колеблется между 16-м и 19-м днями, а затем концентрация прогестерона всегда резко падает. Временной интервал между этим событием и появлением либидо может составить 1-5 дней. Напротив концентрация эстрогенов в периферической плазме крови составляет во время фазы желтого тела полового цикла З.бпг/мл Концентрация эстрогенов остается менее 10 пг/мл" до дня предшествующего охоте, а затем резко возрастает до 15-25 пг/мл и уменьшается за 2-5 часов до начала охоты При определении уровня эстрогена в периферической плазме у группы осемененных коров, но у которых вновь появляется эструс, была отмечена на 3-й день после охоты концентрация, равная 4.1 пг/мл" . на 6-й день после охоты - 6,0 пг/мл На 9-й день отмечен миниматьный, но статистически достоверный пик, который мог отразить волну фолликулярной активности, а также мог вызвать последующую регрессию желтого тела. Пик эстродиола был отмечен от 4-го до 6-го дня после охоты у некоторых животных и, вероятно, был связан с ростом граафовых фолликулов в середине фазы желтого тела. Относительно гормонов гипофиза, у самок характерна цикличность секреции гонадотропина, особенно ярко выделение гонадотропина у половозрелых животных, что проявляется в виде предовуляторного пика ЛГ. Во время фазы желтого тела начальный уровень ЛГ в периферической плазме крови редко превышает 2-3 нг/мл", а во время охоты в крови всех животных наблюдается возрастание уровней, хотя подъем незначительный, пока плазменная концентрация прогестерона снижается ниже 1 нг/мл" . Предовуляторный пик явно соответствует близкому началу охоты (1+8 ч) продолжается обычно не более 6-8 ч, в это время титр ЛГ может увеличиваться до 10-15 нг/мл" и значительно больше (некоторые авторы отмечают пиковые уровни, равные 19-35 нг/мл" и даже - 50-65 нг/мл").
Половой цикл овцы, как было отмечено ранее, сезонный и длится в течение 4-5 месяцев в году. В случае, если овца не осеменена, полицикличность повторяется каждые 16-17 дней, иногда 14-19 дней.
Продолжительность фазы желтого тела составляет не многим более 14 дней. В это время один или несколько предовуляционных фолликулов быстро созревают, достигая диаметра 10-12 мм. Продолжительность охоты бывает 24-30 часов, иногда и больше, овуляция происходит к концу этого периода. т.е. через 24-26 часов после предовуляторной волны секреции гонадотропина. Многие исследователи отмечают, что спаривание овцематок в начале охоты сокращает ожидаемую длительность восприимчивости к самцу: в самом деле, контакт с бараном перед охотой может ускорить предовуляторную волну секреции гонадотропина и. таким образом, сокращать период охоты. Поскольку основные особенности эндокринных изменений, лежащих в основе полового цикла у овцы, во многом схожи с особенностями изменений, происходящих у коров, опишем коротко основные детали. Концентрация прогестерона в периферической плазме в течение первых нескольких дней полового цикла низкая; с 3-11-го 3-11 –ого дня она быстро возрастает "и достигает максимального уровня, равного около 3 нг/мл , что составляет приблизительно половину величины, характерной для крупного рогатого скота в этой же стадии. Для проявления полностью функции желтого тела необходимо наличие как Л1, так и пролактина. Во время фазы желтого тела снова проявляются волны роста фолликулов, которые примерно на 14-й день дают характерный для нее пик фолликулярной фазы. Еще одна особенность, которая отличает половой цикл овцы от цикла коровы, состоит в том, что пик уровня плазменного ЛГ несколько выше (20-100 нг/мл" ) у овец. Этот подъем концентрации гонадотропина не наблюдается до тех пор, пока уровень прогестерона не составит менее 1 нг/мл"
Длительность полового цикла у свиней обычно варьирует в пределах 18-24 дней. Поскольку домашняя свинья полициклична, половой цикл повторяется в течение всего года в отсутствии супоросности, лактации или тяжелых форм стресса, вызванных условиями кормления и содержания. Половая охота длится 40-70 часов, более продолжительна она у свиноматок, чем у ремонтных свинок. Овуляция происходит на 2-й день охоты. Желтое тело функционирует 15-16 дней, во время фазы желтого тела обычно не требуется секреции гипофиза, не считая остаточных количеств предовуляторной волны гонадотропина. Однако, в других исследованиях высказаны предположения, что ткани желтого тела в этот период еще не полностью завершили свой рост и, по-видимому, им требуется трофическая поддержка гормонов гипофиза, как подтвердили опыты с гипофизэктомированными животными.
Фолликулярная фаза цикла длится 5-6 дней, что дольше, чем у коров и овец. В течение этого периода граафовы фолликулы увеличиваются в диаметре от 4-5 мм на 15-й день до 9-11 мм на 21 день. Фактическое число фолликулов, достигающих конечного диаметра в течение одного из циклов, уменьшается к овуляционной норме, характерной для породы или линии животных и, как правило, определяется в начале эструса. Отбор фолликулов, таким образом, происходит в эту последнюю фазу цикла, хотя данные экспериментов на самках с суперовуляцией говорят о том, что это может произойти только благодаря конкуренции циркулирования гонадотропных гормонов. Концентрация i эстрогенов в периферической крови увеличивается от 10-30 пг/мл" до 60-90 пг/мл в течение этого 6-дневного периода созревания фолликулов. В предовуляторный пик концентрация ЛГ в сыворотге может достигать 4-5 нг/мл" . Обычно через 40-48 часов после этого пика происходит овуляция или через 40-42 часа после инъекции препарата ЛГ, сделанной незадолго до эструса, чтобы стимулировать эндогенное выделение гонадотропина.
Миниатюрные породы и линии свиней по времени неодинаково реагируют по подобную инъекцию, что находит отражение в работах ряда авторов. Данные о том, что овуляция наступает через 35-36 часов после начала охоты, незначительно отличаются от вышеназванной величины, составляющей 40-42 часа и, вероятно, зависят от ошибки при определении начала охоты. Концентрация прогестерона увеличивается в предовуляторную фазу от 1,0 нг/мл" к середине фазы желтого тела до 20-35 нг/мл" плазмы перед началом резкого ее уменьшения на 15-16 день цикла
авторсккие права: .Л. К. Эрнст, А. Н. Варнавский